1. Синдром ломкой хромосомы X. Синдром ломкой X хромосомы. Признаки синдрома ломкой X хромосомы.
2. Наследование сцепленное с Y хромосомой. Y - сцепленные заболевания. Y-сцепленное наследование. Наследование сцепленное по половой хромосоме Y.
3. Пол человека. Генетические методы определения пола человека. Генетические механизмы определения пола.
4. Дифференцировка пола человека. Гены дифференцировки пола человека. Экспрессия генов, определяющих пол плода.
Дифференцировка пола человека. Гены дифференцировки пола человека. Экспрессия генов, определяющих пол плода.
Ген SRY экспрессируется в половом гребешке в момент индукции дифференциации яичек. В случае делеций или иных мутаций в гене SRY яички не дифференцируются. Продукт гена SRY является, вероятно, ядерным фактором транскрипции, связывая белки, модулирующие транскрипцию. Белок SRY содержит HMG-бокс, который связывается с кон-сенсусной последовательностью ААТААЦ ДНК. Примерно у 20 % 46,ХУ-мужчин ген SRY yt обнаруживают, что позволяет предположить его неабсолютную обязательность для мужской дифференциации пола.
Установлено, что, кроме гена SRY, несколько аутосомных генов играют определенную роль в мужской дифференциации пола. Один из них — ген супрессор опухоли Вильмса, кодирует фактор транскрипции, связывающийся со специфическими ДНК-последовательностями. Точковые мутации в этом гене иногда приводят к формированию гонадно противоположного пола. Второй ген такого же типа — SOX9. Белок, кодируемый этим геном, также содержит HMG-бокс. Мутации в гене SOX9 вызывают кампомелическую диспла-зию, при которой у индивидуумов 46, XY-фенотипически развивается женский пол.
Стероидогенный фактор 1 (SF1), кодируемый соответствующим геном, является фактором транскрипции, регулирующим экспрессию ряда генов, которые участвуют в продукции стероидных гормонов и мужской половой дифференциации. Показано, что гемизиготная мутация в этом гене ассоциирует с надпочечниковой недостаточностью и гонадным дисгенезом у лиц с кариотипом 46,XY. Ген MIS, картированный в области 19р13, отвечает за синтез фактора, ингибирующего парамезонефральные протоки. Мутации в этом гене ведут к проявлениям синдрома персистен-ции указанных протоков. Экспрессия MIS регулируется продуктами генов S0X9 и SF1.
Для нормального развития гонад необходимы также X-хромосомные локусы. У некоторых лиц с гонадным дисгенезом 46,XY наблюдалась дупликация области Хр21. В этой области находится ген DAX1, белок которого функционирует как транскрипционный фактор. Еще один ген в хромосоме X — ХН2, играет не вполне выясненную роль в дифференцировке яичек. Он кодирует фермент геликазу, которая расплетает ДНК, делая ее доступной для транскрипционных факторов.
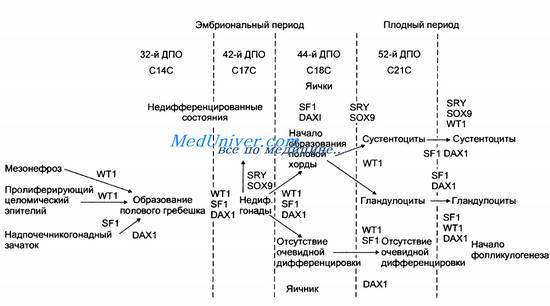
Нет сомнений в том, что современные представления о генетическом контроле становления пола далеко не полные. Это относится не только к тому, какие гены вовлечены в данный процесс, но и к тому, на каких стадиях развития и в каких тканях эти гены проявляются. На примере генетического контроля определения пола мы сталкиваемся с некоторыми кардинальными проблемами генетики развития — ткане- и времяспецифичности действия генов. При образовании полового гребешка из мезонефроса и целомического эпителия в соответствующих тканях обнаружена активность гена WT1, а в зачатке надпочечника, который также имеет отношение к образованию полового гребешка, — генов SF1 и DAX1. Половой гребешок формирует недифференцированные гонады при участии тех же генов.
В предшественниках половых клеток эмбриона мужского пола резко возрастает активность генов SRY и SOX9. Практически все перечисленные гены и еще ген MIS активны в сустентоцитах, а гены SF1 и DAX1 — в гландулоцитах. В то же время у женбкого эмбриона вплоть до плодного периода гонады остаются недифференцированными, хотя в них обнаруживают активность генов WT1, SF1 и DAX1 (рис. 5.8).
- Вернуться в оглавление раздела "Генетика. Медицинская генетика."