Консервативность рецепторных систем. Избыточность рецепторов
Важным свойством рецепторно-ферментных систем является их консервативность в отношении концентраций ряда параметров. Выше мы уже говорили о различных механизмах инактивации и регуляции системы усиления сигнала, приводящих в конце концов к возвращению концентрации вторичных посредников к исходной. Однако консервативность рецепторно-ферментных систем выходит за рамки поддержания одной клеткой концентраций ряда метаболитов.
Изучение рецепторно-ферментных систем одного типа у людей и отдельных животных позволило установить наличие консервативности совсем другого рода.
1. Исследование параметров опиоидных рецепторов мозга отдельных животных, подвергшихся наркотизации (животным длительное время вводили морфин), показало, что при значительной вариации индивидуальных параметров [R]o и К отмечалось относительное постоянство (с точностью до 30%) их отношения [R]o/K (Зайцев с соавт., 1986а).
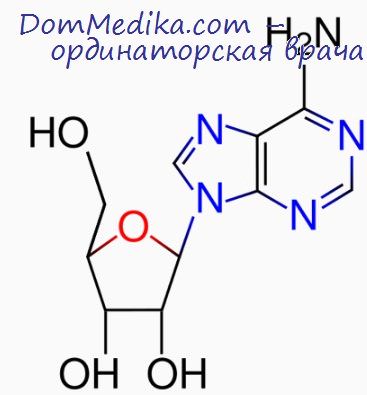
2. Проведенное нами (Шевченко с соавт., 1988) исследование количественных зависимостей скорости и степени агрегации тромбоцитов от концентрации PGE показало, что значения IC50 (концентрации PGEi, при которой наблюдается 50%-е ингибирование агрегации) для тромбоцитов разных индивидуумов различны: в наших экспериментах они изменялись в пределах 17-100 нМ PGE1. Значения параметров связывания PGE1 с рецепторами тромбоцитов тоже резко отличались. Однако число молекул PGEX. специфически связавшихся с одним тромбоцитом при концентрации PGE1, равной IC50, оказалось практически постоянной величиной для тромбоцитов разных индивидуумов: 10±2 молекулы PGEi на тромбоцит.
Полученные результаты можно объяснить, если полагать, что несмотря на различия параметров [R]o и К у разных людей или отдельных животных, количественные характеристики пострецепторных звеньев клеточного ответа консервативны, т.е. мало отличаются друг от друга внутри данной группы индивидуумов или данной группы животных. При этом поскольку для опиоидных рецепторов животных имеет место феномен "резерва" рецепторов (Chavkin, Goldstein, 1984), то, как уже говорилось в предыдущем разделе, клеточный ответ определяется отношением [R]0/K. Следовательно, при постоянстве этого параметра у различных животных при данной фиксированной дозе наркотического соединения степень заселенности опиоидных рецепторов у разных животных будет близкой. С другой стороны, "избыточные" рецепторы для PGE на тромбоцитах отсутствуют (Шевченко с соавт., 1988).
Следовательно, в етом случае заселенность рецепторов определяется как их концентрацией, так и сродством к лиганду. Помимо этого, как мы видим, в тромбоцитах имеет место адаптация рецепторного аппарата человека к индивидуальной концентрации простаноида.
- Читать далее "Дискриминация моделей процессов комплексообразования. Графические представления взаимодействия рецепторов"
Оглавление темы "Рецепторы центральной нервной системы":1. Рецепторно-ферментные системы. Рецепторы как датчики сигнала
2. Константы скоростей нейрогормонов. Гетерогенность популяции рецепторов
3. Усиление сигнала рецепторами. Ферментативное усиление сигнала
4. Каскадные системы усиления сигнала. Даун регуляция рецепторов
5. Инактивация лиганд-рецепторного комплекса. Двойное действие лиганда на рецепторы
6. Суперчувствительность рецепторов. Кооперативность рецепторов
7. Механизмы суперчувствительности рецепторов. Суицидные ферментные комплексы
8. Значение концентрации лиганда. Рецепторный резерв
9. Консервативность рецепторных систем. Избыточность рецепторов
10. Дискриминация моделей процессов комплексообразования. Графические представления взаимодействия рецепторов