Белок APOBEC3G как механизм резистентности к ВИЧ
Несколько лет назад Малим (Malim) и соавторы описали потенциальную функцию вирусного белка Vif, которая состоит в нейтрализации некоей внутриклеточной активности, направленной против ВИЧ в клетках человека.
Устойчивость клетки к прогрессированию вируса в ней, по-видимому, связана с этапами цикла репликации вируса, на которые может действовать какой-то врожденный внутриклеточный фактор. Эти данные были получены во время исследований, в которых проводилось сравнение мутантных разновидностей ВИЧ-1, у которых отсутствует ген vif с ВИЧ-1 дикого типа, содержащих ген vif. В этих опытах было показано, что некоторые Т-клетки (например, клеточной линии CEM-SS) вполне пермиссивны в отношении поддержания репликации обоих вирусов, но большинство других Т-клеток, например линии HUT 78, а также свежевыделенные мононуклеарные клетки из периферической крови, не поддерживали инфекцию вирусам, не содержащими Vif.
В клетках с делецией гена vif интенсивность репликации вируса была в 50 раз ниже. По-видимому, белок Vif взаимодействует с неким клеточным белком на ранних стадиях инфекционного процесса, что обеспечивает более эффективное размножение вируса в клетке.
Дальнейшие исследования привели к открытию в клетках человека гена, первоначально названного СЕМ-15, который тормозил репликацию ВИЧ; эффект присутствия этого гена отменялся действием белка Vif. Впоследствии было показано, что СЕМ-15 представляет собой известный ген человека, кодирующий фермент, катализирующий «редактирование» информационной РНК для аполипопротеина В, каталитическую полипептидподобную 3G молекулу (APOBEC3G - apolipoprotein В mRNA editing enzyme, catalytic polypertide-like 3G), который осуществляет дезаминирование ДНК.
APOBEC3G вызывает появление мутаций с заменой G на А во вновь синтезируемой вирусной ДНК. Под действием этого белка происходит замена цитозина на урацил в ретровирусной минус-цепи ДНК. В результате таких замен происходит повышение частоты мутаций типа «замена G на А» в плюс-цепи ДНК.
Инактивация вирусного потомства происходит как в результате подобных мутаций, так и за счет деградации ДНК, которая инициируется под действием вирусных N-гликозидаз и щелочных нуклеаз. Vif, взаимодействуя с APOBEC3G, блокирует его активность и тем самым предотвращает накопление фатальных генетических мутаций в вирусном потомстве, чем обеспечивает продукцию инфекционно дееспособного «здорового» ВИЧ.
Каталитический домен APOBEC3G играет важную роль в его функционировании, а цинк-связывающий домен необходим для дезаминирования цитидина и противовирусного действия. Другой домен (CD1) отвечает за связывание с РНК и включение APOBEC3G в вирусные частицы.
Неизвестно, насколько инкорпорация APOBEC3G в вирионы зависит от РНК вирусного генома, но нуклеокапсидный домен Gag-белка ВИЧ для этого процесса необходим.
Vif вызывает убиквитинилирование и распад APOBEC3G под действием клеточных белков Cullen5 (CUL5), элонгенов b и с, и RBX1. По-видимому, в обеспечении этих функций участвуют два различных домена Vif: один, связывающий APOBEC3G, а другой, вызывающий его деградацию в протеосомах.
Vif также предотвращает включение APOBEC3G в вирионы и, таким образом, инфицирование новых клеток дочерними вирусами может проходить без дезаминирования. Геномная РНК вируса помогает стабилизировать связь APOBEC3G с ядерным белковым комплексом ВИЧ-1 внутри вириона. После того, как APOBEC3G включается в вирионы, эффективность торможения его противовирусной активности со стороны Vif может снижаться.
Сходное с APOBEC3G противовирусное действие было отмечено у другого клеточного белка - APOBEC3F, и Vif также может ослаблять его действие. Совсем недавно было показано, что еще один член семейства АРОВЕС, APOBEC3DE может подавлять инфекции ВИЧ-1 и SIV за счет дезаминирования цити-дина в некоторых новых специфических участках провирусных ДНК.
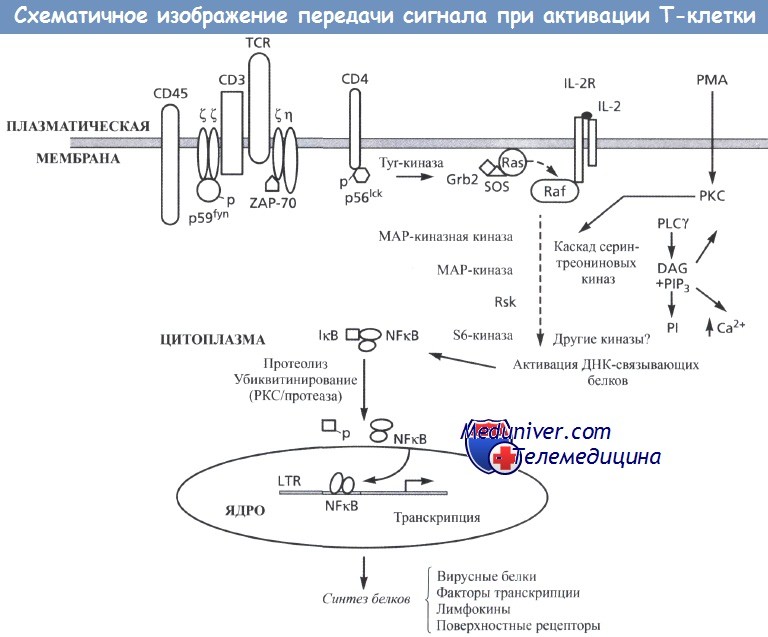
Активация начинается с генерации сигнала на плазматической мембране (верхняя часть рисунка),
опосредованной взаимодействием рецептора с лигандом, например рецептора интерлейкина-2 (IL-2R) + IL-2,
взаимодействием рецептора Т-лимфоцитов для антигена (TCR) + CD4 или поступлением молекул, способных проникать через мембрану (например, форбол миристат ацетат [РМА]).
В случае с TCR стимуляция приводит к активации тирозинкиназ (верхняя левая часть рисунка), в частности Fyn (p59fyn), ZAP-70 и Lck (p56lck).
Эти процессы, возможно, регулируется тирозин-фосфатазами, например молекулой CD45.
Активированные тирозинкиназы фосфорилируют различные цитоплазматические субстраты, которые инициируют цепь процессов по проведению сигнала.
Например, Ras (верхняя средняя часть рисунка) может активироваться после связывания с Grb2 и SOS. Это приводит к активации цитоплазматических сериновых киназ, например Raf, что вызывает активацию каскада серин-треониновых киназ (например, МАР-киназной киназы, МАР-киназы, Rsk и S6-киназ), таким образом усиливая активационный сигнал.
Каскад серин-треониновых киназ приводит к активации ДНК-связывающих белков и факторов транскрипции, которые регулируют экспрессию генов. Фосфорилирование и/или протеолиз IкВ в цитоплазме (нижняя левая часть рисунка) приводит к его диссоциации от NF-кВ и позволяет NF-кВ проникать через ядерную мембрану, а затем связываться с ДНК и активировать транскрипцию и синтез белка. Такой процесс трансдукции сигнала также может инициироваться при активации протеинкиназы С (РКС) эфирами форбола (например, РМА) (верхняя правая часть рисунка).
Активированная РКС вызывает активацию фосфолипазы Су (PLCy) с образованием вторичных мессенджеров - диацилглицерола (DAG) и фосфоинозитолтрифосфата (PIP3), что приводит к мобилизации внутриклеточного кальция (Са2+) и высвобождению фосфоинозитола (PI).
- Читать "Аналоги белка APOBEC3G у животных"
Оглавление темы "Репликация ВИЧ":- Этапы репликации ВИЧ внутри клетки
- Влияние внутриклеточных факторов на репликацию ВИЧ
- Особенности репликации ВИЧ в Т-клетках
- Особенности репликации ВИЧ в моноцитах и макрофагах
- Белок APOBEC3G как механизм резистентности к ВИЧ
- Аналоги белка APOBEC3G у животных
- Природные функции белков APOBEC
- Белок TRIM5a как механизм резистентности к ВИЧ
- Влияние цитокинов на репликацию ВИЧ
- Влияние белков ВИЧ на клетку