Этапы инфицирования CD4 клеток ВИЧ-инфекцией
Исследования, проведенные вскоре после первого выделения ВИЧ, показали, что CD4+-Т-лимфоциты являются очевидными клетками-мишенями для ВИЧ-инфекции. В лимфатических тканях можно обнаружить до 100 миллионов инфицированных покоящихся CD4+-Т-клеток памяти (HLA-DR- CD45RO+) и около 5 миллиардов вирионов в составе иммунных комплексов, связанных с фолликулярными дендритными клетками стромы. Кроме того, необходимо учитывать наличие большое количества инфицированных CD4+-Т-клеток памяти в лимфоидных тканях кишечника.
При исследовании относительной чувствительности активированных CD4+-клеток к ВИЧ-инфекции обнаруживают значительную вариабельность, не связанную с продукцией хемокинов. Завершившие лимфопоэз неиммунные (то есть до контактов с антигеном и до вступления в иммунный ответ) CD4+-Т-лимфоциты обычно находятся в неактивной форме (HLA-DR- CD38+ CD45RA).
Во время острой инфекции они активируются и выполняют функции лимфоцитов-эффекторов и CD4+-клеток памяти (HLA-DR+ CD38hi CD45RO).
Некоторые активированные CD4+-клетки-памяти могут возвращаться в состояние покоя, то есть превращаться в центральные клетки-памяти (HLA-DR- CD69"+ CD45RO). Как активированные, так и покоящиеся (CD45RO) CD4+ клетки чувствительны к ВИЧ-инфицированию in vivo (см. ниже). Несмотря на то, что тимус, очевидно, не является основным местом развития ВИЧ-инфекции, провирус ВИЧ можно обнаружить в неиммунных CD4+ CD45RA+ CD62+ Т-клетках.
Это может зависеть от цитокинового окружения в тимусе. Некоторые цитокины, в частности IL-4, IL-6, IL-7 и IL-15, могут повышать восприимчивость неиммунных CD4+-Т-клеток к ВИЧ-инфекции. Однако применение препарата IL-2 в качестве терапевтического средства существенно не изменяет чувствительность неиммунных Т-лимфоцитов (CD45RA+ к продуктивной ВИЧ-инфекции, но предварительное воздействие препаратом IL-7 увеличивает продуктивное инфицирование в 58 раз.
IL-7 усиливает поверхностную экспрессию CXCR4, но не CCR5, и приводит к интенсификации пролиферации неиммунных Т-клеток.
In vivo активированные CD4+-клетки производят самые большие количества инфекционного вируса, однако концентрация вируса также зависит и от пусть не столь активной репликации ВИЧ в покоящихся CD4+-клетках памяти. Инфицированные активированные CD4+-Т-клетки могут производить до пяти раз больше вирионов, чем инфицированные покоящиеся клетки памяти.
Важно отметить, что >90% продуктивно инфицированных клеток при острой инфекции представляют собой покоящиеся CD4+-T-клетки памяти, и скорость обновления этих клеток может занимать до 14 дней; время полужизни инфицированной активированной CD4+-клетки составляет около одного дня. Таким образом, концентрация вируса в плазме крови будет преимущественно определяться продукцией вируса в течение длительного времени покоящимися CD4+-клетками памяти.

ВИЧ-специфичные CD4+-T-клетки памяти также содержат большее количество провирусной ДНК ВИЧ, чем CD4+ клетки памяти с другой специфичностью антигенраспознающего рецептора на всех стадиях ВИЧ-инфекции. Данные наблюдения четко указывают на повышенную чувствительность этих клеток к инфицированию и, следовательно, разрушению. Активированные, полностью дифференцированные CD4+-Т-клетки, которые обладают слабой пролиферативной активностью, содержат в 10 раз меньше копий провирусной ДНК ВИЧ.
Некоторые инфицированные клетки несут латентный интегрированный провирус; в других геном вируса находится в преинтегрированном состоянии. Клетки, содержащие интегрированный ВИЧ, представляют часть существующего длительное время резервуара инфекции в организме и могут сохраняться до четырех лет.
Что касается экспрессии рецепторов для хемокинов, неиммунные (CD45RA) CD4-Т-лимфоциты, также как и CD4+-клетки-памяти, экспрессируют рецептор CXCR4, тогда как наличие CCR5, по-видимому, характерно преимущественно для субпопуляции активированных и покоящихся CD45RO-T-клеток памяти. Таким образом, в норме гораздо меньшее количество клеток экспрессирует CCR5, чем CXCR4.
Эти данные говорят о том, что вирусы биотипа R5 могут заражать только CCR5-экспрессирующие Т-клетки, тогда как вирусы биотипа Х4 могут инфицировать любые CD4+-Т-клетки, в том числе и неиммунные, имеющие рецептор CXCR4. Несмотря на то, что окончательный вывод делать еще рано, эти данные могут объяснять нарастающую гибель CD4+-клеток после появления Х4-вируса в организме хозяина.
Покоящиеся CD4+-клетки памяти, по-видимому, подвергаются инфицированию в первую очередь и преимущественно R5-вирусами. Такое преимущественное инфицирование ВИЧ-1 может быть связано с экспрессией молекулы интегрина LFA-1 на этих клетках. С этим может быть связано преобладание R5-вирусов на ранних стадиях инфицирования. Кроме того, повышенная способность R5-вирусов к репликации в пермиссивных CD4+ CCR5+ CXCR4+ Т-клетках может также объяснять присутствие R5-биотипа во время острой или первичной инфекции.
Также было показано, что продукция инфекционных вирусов биотипа R5 на одну CCR5+-клетку-мишень в пять-десять раз выше, чем продукция вирусов биотипа Х4 ВИЧ-1. Однако в других исследованиях, проведенных с использованием клонов вирусов, не наблюдали различий в репликации между вирусами биотипов R5 и Х4. Возможно, более высокий уровень репликации R5-вирусов связан с продукцией вируса, обладающего менее выраженными цитопатогенными свойствами.
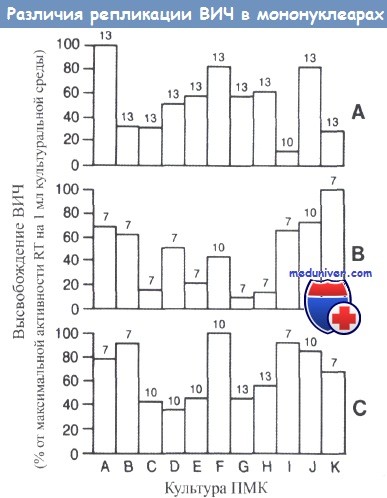
Клетки, полученные от 11 различных доноров азиатского (столбики от А до D), афроамериканского (столбики Е, F и G) и европейского (столбики от Н до К) происхождения были инфицированы тремя различными штаммами ВИЧ-1 (SF2 [A], SF247 [В] и SF170 [С]).
Наблюдали воспроизводимые различия в интенсивности репликации и во времени максимального высвобождения вируса (число над столбиком).
CD4+-Т-лимфоциты, различающиеся по профилю экспрессируемых цитокинов (типа Тh-1 или Th-2), могут также различаться и по профилю экспрессируемых рецепторов к хемокинам и, следовательно, по чувствительности к инфицированию ВИЧ-1. Рецепторы CXCR3 и CCR5 преимущественно экспрессируются на Th1-клетках человека. Th2 CD4+-клетки экспрессируют рецепторы CXCR4, CCR3, и CCR5.
Два типа Th одинаково экспрессируют рецептор CXCR4; однако было показано, что экспрессия CCR5 в 8 раз выше в Thl-клетках по сравнению с Тh2. Интенсивность репликации ВИЧ также в 4 раза выше в субпопуляции Тh1 по сравнению с Тh2. Однако другие исследования показали прямо противоположный результат, а именно, репликация как R5, так и Х4-вирусов в Тh2-клетках была активнее, чем в Тh1-клетках.
В некоторых работах было показано, что R5-изоляты, очевидно, инфицировали все типы CD4+-лимфоцитов, тогда как Х4-изоляты ВИЧ размножались преимущественно в клетках Тh2-типа. Этот результат был неожиданным, поскольку, как отмечали выше, все циркулирующие неиммунные Т-клетки экспрессируют рецептор CXCR4, но только активированные Т-клетки памяти экспрессируют рецептор CCR5. В других исследованиях оба типа клеток проявляли одинаковую чувствительность к ВИЧ-инфекции, но интенсивность репликации вируса в Тh1 клетках была несколько выше.
При использовании в исследованиях нестимулированных мононуклеарных клеток из пуповинной крови или клонов CD4+-клеток некоторые ученые обнаружили, что в них преимущественно реплицируются изоляты R5, а не Х4. Для зрелых мононуклеарных клеток (то есть из периферической крови взрослых) наблюдают обратную картину. Напротив, изоляты Х4 проявляют большую способность к инфицированию первичных CD4+-лимфоцитов, чем изоляты R5. Некоторые исследования позволили предположить, что ВИЧ-1 реплицируется в 3 раза активнее в Т-лимфоцитах пуповинной крови, чем в лимфоцитах крови взрослого человека, и в 9 раз активнее в макрофагах, образовавшихся из моноцитов, пуповинной крови, чем в макрофагах взрослого человека. Эффект, по-видимому, связан с увеличением LTR-зависимой транскрипции ВИЧ-1.
Некоторые различия между CD4+-клетками по чувствительности к инфицированию, особенно отмечавшиеся в ранних исследованиях, скорее всего, связаны с особенностями использованного штамма вируса. Способность определенных цитокинов модулировать экспрессию хемокиновых рецепторов (например, IL-2 и IL-7) может также влиять на чувствительность CD4+-клеток к ВИЧ-инфекции. Некоторые исследования также показали способность вирусов биотипа R5 активировать CD4+-Т-клетки и тем самым способствовать увеличению интенсивности репликации вируса в этих клетках.
Этот процесс может протекать с участием механизмов внутриклеточной передачи сигнала от рецептора к хемокинам CCR5. Это может оказывать относительное влияние на импорт преинтеграционного комплекса в ядро клетки. Например, импорт в ядро провирусной ДНК вирусов биотипа R5 не зависит от активации CD4+-Т-клеток через рецепторы CD3/CD28. Эти результаты помогают объяснить преобладание R5-изoлятoв на ранних стадиях инфекции и гибель клеток Th1-типа по мере прогрессирования заболевания у пациентов, если не происходит смены биотипа вируса.
Также были описаны различия по интенсивности продукции вируса в мононуклеарных клетках периферической крови (среди которых основными клетками-мишенями являются CD4+-лимфоциты) у различных людей. Штаммы ВИЧ-1 или ВИЧ-2 могут размножаться до высоких титров в этих клетках, полученных только от некоторых людей. В ряде случаев были продемонстрированы тысячекратные различия в интенсивности репликации вируса. Не было найдено ни одного штамма ВИЧ, который бы одинаково хорошо размножался в мононуклеарных клетках периферической крови, полученных от любого человека.
Аналогично мононуклеарные клетки периферической крови одного и того же человека различаются по чувствительности к различным изолятам ВИЧ. В отдельных редких случаях, как отмечали выше, мононуклеары крови (как CD4+-лимфоциты, так и моноциты) проявляли устойчивость к инфицированию R5-вирусами, поскольку на клетках отсутствовал ССR5-корецептор. Таким образом, предполагают, что при инфицировании двух человек одним и тем же вирусом последний может активировать различные механизмы патогенеза в зависимости от его способности размножаться и распространяться в периферических лимфоцитах каждого человека.
Подводя итог, можно сказать, что чувствительность CD4+-Т-клеток к заражению и их способность поддерживать репликацию вируса, по-видимому, определяется рядом факторов: свойствами вируса, генетическими свойствами клетки хозяина, которые определяют наличие на ней тех или иных поверхностных молекул, и внеклеточными факторами. Отдельные субпопуляции CD4+-клеток, по-видимому, могут быть устойчивы к инфицированию определенными биотипами ВИЧ.
Факторы, влияющие на ВИЧ-инфицирование и репликацию вируса:
• Наличие клеточных рецепторов для вируса
• Свойства белков оболочки вируса (строение, кон-формация, заряд)
• Степень гликозилирования белков оболочки вируса
• Количество «шипов» на оболочке (степень шеддинга gp120)
• Клеточные протеазы
• Взаимодействие внутриклеточных факторов с вирусной последовательностью нуклеотидов LTR и с вирусными белками (например, Tat, Rev и Net)
• Степень экспрессии вирусных регуляторных и добавочных генов (например tat, rev, nef, vif, vpu и vpr)
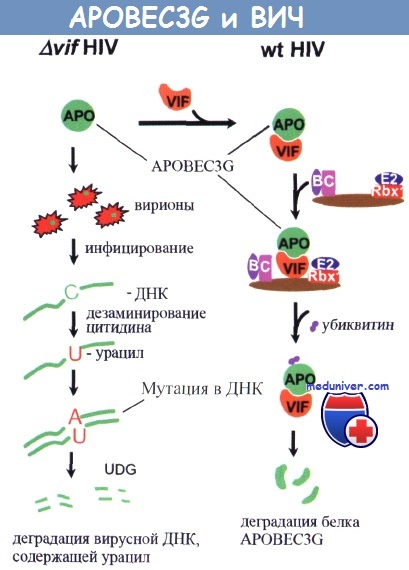
APOBEC3G включается в состав вирионов в отсутствие Vif-белка.
После инфицирования APOBEC3G вызывает дезаминирование цитидина, превращая его в урацил (С=>U) на минус-нити вирусной ДНК во время обратной транскрипции С=>U преобразуется в мутацию G=>А плюс-цепи во время синтеза плюс-цепи.
Урацилсодержащая вирусная ДНК разрушается под действием клеточных ферментов.
Vif предотвращает APOBEC3G включение клеточного белка в вирионы, вызывая убиквитинзависимое расщепление APOBEC3G.
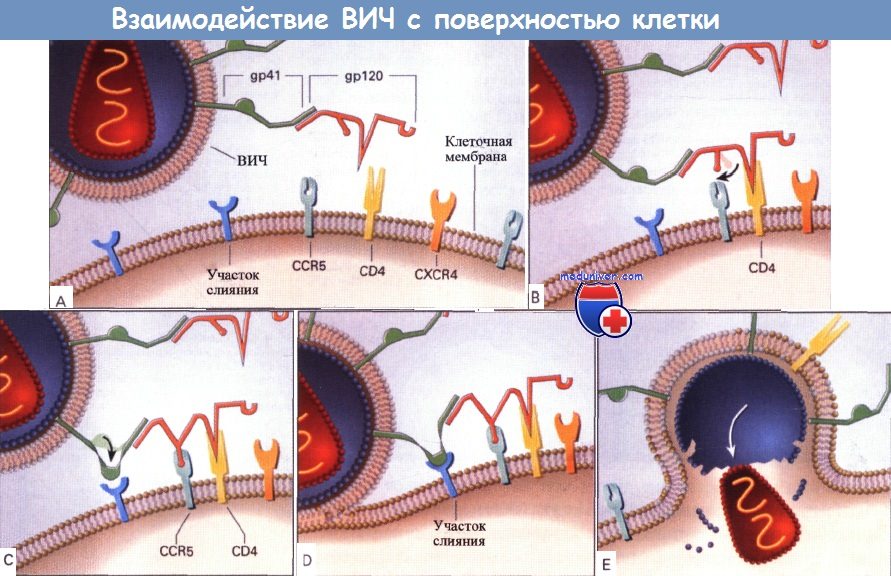
(А) ВИЧ взаимодействует с рецептором на поверхности клетки, обычно с CD4, и за счет конформационных изменений более плотно связывается с клеткой, взаимодействуя с другими поверхностными молекулами,
например с хемокиновыми рецепторами CXCR4 и CCR5. В качестве альтернативы некоторые вирусы, например определенные штаммы ВИЧ-2, могут непосредственно связываться с CXCR4.
(от В до Е) Вероятно, ВИЧ-инфицирование проходит через следующие основные этапы. Центр связывания CD4 на gp120 ВИЧ-1 взаимодействует с молекулой CD4 на поверхности клетки.
Конформационные изменения вирусной оболочки и CD4-рецептора обеспечивают связывание gp120 с другим поверхностным рецептором клетки, например с CCR5.
За счет этого взаимодействия вирусная оболочка приближается к поверхности клетки, что приводит к взаимодействию gp41 вирусной оболочки с доменом слияния на поверхности клетки.
ВИЧ сливается с клеткой. Затем вирусный нуклеоид проникает в клетку, скорее всего, с помощью других клеточных механизмов. Сразу после этого начинается репликация вируса.
- Вернуться в оглавление раздела "Инфекционные болезни"
Оглавление темы "Механизмы ВИЧ-инфекции":- Клиника острого ВИЧ-инфицирования
- Лабораторная диагностика острого и первичного ВИЧ-инфицирования
- Иммунологические показатели острого и первичного ВИЧ-инфицирования
- Свойства распространяющегося вируса острого и первичного ВИЧ-инфицирования
- Клеточный иммунный ответ организма на острое ВИЧ-инфицирование
- Образование противовирусных антител на острое ВИЧ-инфицирование
- Обзор патогенеза первичного ВИЧ-инфицирования
- Клетки и ткани инфицируемые ВИЧ
- Этапы инфицирования клеток ВИЧ-инфекцией
- Этапы инфицирования CD4 клеток ВИЧ-инфекцией