Особенности репликации ВИЧ в Т-клетках
Для репликации ВИЧ необходима активация Т-клеток. При этом происходит взаимодействие обычных внутриклеточных факторов транскрипции с вирусными LTR. Существенное значение для активации транскрипции с вирусных генов имеет также вирусный белок Tat.
Такая активация клетки является частью механизма внутриклеточной передачи сигналов от митогенов или антигенов (последние передаются через Т-клеточные рецепторы для антигенов и усиливаются через сигналы от корецепторов, в первую очередь через взаимодействие CD28 - В7), что приводит к усилению экспрессии генов. Заметная костимуляция активации лимфоцитов происходит также через взаимодействие LFA-3 с CD2. Этот процесс вызывает нарастание концентрации свободного кальция внутри клетки и зависит от активации кальцийзависимой протеинкиназы С и других процессов, а именно фосфорилирования белков.
Во время такой стимуляции Т-клеток активируются факторы транскрипции. Например, NF-кВ обычно находится в связанном состоянии со своим ингибитором IкВ. Однако фосфорилирование IkB, катализируемое протеинкиназой С, приводит к диссоциации IkB от NF-kB и к последующей транслокации активного фактора транскрипции NF-kB в ядро, где он связывается с комплементарным участком вирусной LTR в провирусе ВИЧ.
Взаимодействие клеточных факторов транскрипции с вирусными последовательностями нуклеотидов в LTR может усиливать репликацию вируса или подавлять ее (например, YYI). В случае некоторых мутантных вирусов биотипа Х4 было показано, что делеция последовательностей нуклеотидов в ДНК LTR провируса, связывающих клеточный фактор транскрипции NF-kB, приводит к снижению интенсивности репликации ВИЧ. Тем не менее некоторые изоляты ВИЧ-1 и некоторые мутантные варианты SIV, у которых отсутствуют сайты связывания NF-kB на LTR, могут хорошо реплицироваться в мононуклеарных клетках периферической крови.
Кроме того, было показано, что SIV, мутантный по сайту связывания NF-kB, может вызывать СПИД у обезьян. В настоящее время предпринимают попытки идентификации всех клеточных факторов, участвующих в репликативном цикле ВИЧ. Некоторые цитокины и гормоны, а также трансактивирующие белки других вирусов также могут усиливать продукцию ВИЧ, индуцируя или включаясь во внутриклеточные механизмы активации.

После проникновения ВИЧ в CD4+-клетку может происходить перемещение молекул тирозинкиназы р56lck семейства Src от цитоплазматического участка молекулы CD4, что приводит к активации протеинкиназы С и усилению продукции вируса. Поскольку тирозинкиназа р56lck, по-видимому, действует на различные внутриклеточные мишени, в частности, на рецептор для интерлейкина-2 (IL-2), она может прямо или опосредованно воздействовать на активность клеточных факторов, связывающихся с LTR ВИЧ.
В некоторых ранних исследованиях было показано, что связывание нативного gp120 с CD4 само по себе вызывает активацию CD4+-лимфоцитов, которая выражается в увеличении внутриклеточных концентраций инозитол трифосфата и кальция, а также в индукции экспрессии рецептора для IL-2. Это наблюдение, однако, не было подтверждено другими исследователями, которые сообщали об отсутствии влияния связывания ВИЧ на фосфорилирование CD4,4 CD4-зависимую активность киназы p56lck или поступления кальция в клетку.
К тому же было показано, что блокатор кальциевых каналов верапамил не влияет на проникновение ВИЧ в клетку. Таким образом, роль повышения концентрации внутриклеточного кальция на начальных этапах проникновения ВИЧ в Т-клетки до конца не определена. Тем не менее, gp120 может вызывать дополнительное поступление кальция в клетку в определенных линиях перевивных нейрональных клеток с помощью механизма, который блокируется верапамилом, но не связан с участием молекулы CD4.
То есть, «сигнал-линг» через молекулу CD4, по-видимому, не является необходимым для продукции ВИЧ-инфицированной клеткой.
Необходимо отметить, что в некоторых исследованиях наблюдали блокировку репликации ВИЧ-1 биотипа R5 в CD4+-клетках памяти, но репликация ВИЧ-1 биотипа Х4 в таких клетках была возможна. Механизмы блокировки размножения вирусов R5 в клетках памяти неизвестны, но такая возможность может объяснять более низкую патогенность R5-вирусов для CD4+-Т-клеток памяти. Кроме того, покоящиеся CD4-клетки, по-видимому, проявляют устойчивость к ВИЧ-инфекции еще и за счет действия внутриклеточного белка APOBEC3G.
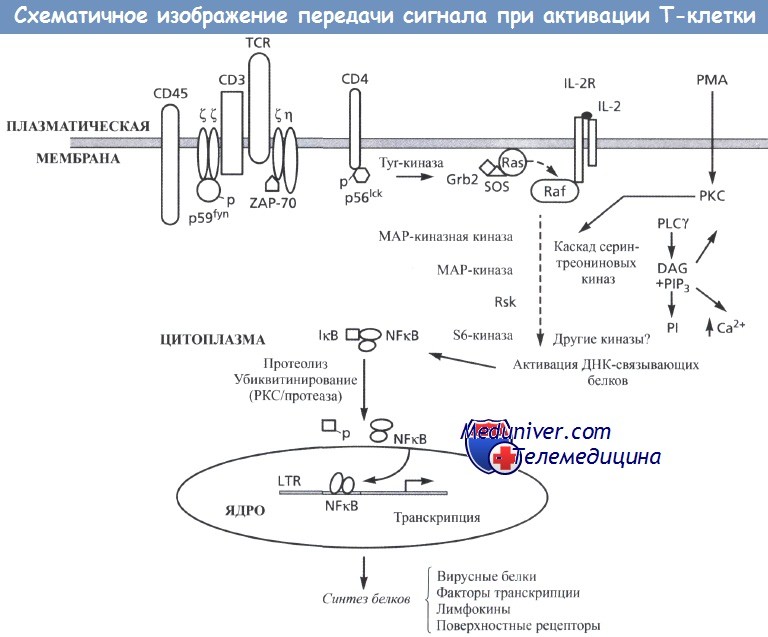
Активация начинается с генерации сигнала на плазматической мембране (верхняя часть рисунка),
опосредованной взаимодействием рецептора с лигандом, например рецептора интерлейкина-2 (IL-2R) + IL-2,
взаимодействием рецептора Т-лимфоцитов для антигена (TCR) + CD4 или поступлением молекул, способных проникать через мембрану (например, форбол миристат ацетат [РМА]).
В случае с TCR стимуляция приводит к активации тирозинкиназ (верхняя левая часть рисунка), в частности Fyn (p59fyn), ZAP-70 и Lck (p56lck).
Эти процессы, возможно, регулируется тирозин-фосфатазами, например молекулой CD45.
Активированные тирозинкиназы фосфорилируют различные цитоплазматические субстраты, которые инициируют цепь процессов по проведению сигнала.
Например, Ras (верхняя средняя часть рисунка) может активироваться после связывания с Grb2 и SOS. Это приводит к активации цитоплазматических сериновых киназ, например Raf, что вызывает активацию каскада серин-треониновых киназ (например, МАР-киназной киназы, МАР-киназы, Rsk и S6-киназ), таким образом усиливая активационный сигнал.
Каскад серин-треониновых киназ приводит к активации ДНК-связывающих белков и факторов транскрипции, которые регулируют экспрессию генов. Фосфорилирование и/или протеолиз IкВ в цитоплазме (нижняя левая часть рисунка) приводит к его диссоциации от NF-кВ и позволяет NF-кВ проникать через ядерную мембрану, а затем связываться с ДНК и активировать транскрипцию и синтез белка. Такой процесс трансдукции сигнала также может инициироваться при активации протеинкиназы С (РКС) эфирами форбола (например, РМА) (верхняя правая часть рисунка).
Активированная РКС вызывает активацию фосфолипазы Су (PLCy) с образованием вторичных мессенджеров - диацилглицерола (DAG) и фосфоинозитолтрифосфата (PIP3), что приводит к мобилизации внутриклеточного кальция (Са2+) и высвобождению фосфоинозитола (PI).
- Читать "Особенности репликации ВИЧ в моноцитах и макрофагах"
Оглавление темы "Репликация ВИЧ":- Этапы репликации ВИЧ внутри клетки
- Влияние внутриклеточных факторов на репликацию ВИЧ
- Особенности репликации ВИЧ в Т-клетках
- Особенности репликации ВИЧ в моноцитах и макрофагах
- Белок APOBEC3G как механизм резистентности к ВИЧ
- Аналоги белка APOBEC3G у животных
- Природные функции белков APOBEC
- Белок TRIM5a как механизм резистентности к ВИЧ
- Влияние цитокинов на репликацию ВИЧ
- Влияние белков ВИЧ на клетку
- Как поддерживать здоровье при гепатите: что можно и нельзя