Механизм слияния клеток в синцитий при ВИЧ
В слиянии клеток могут играть роль другие участки молекул CD4 и белков вирусной оболочки (например, gp10), чем те, что участвуют в прикреплении вируса к клетке. Некоторые моноклональные антитела к CD4 не блокируют связывание вируса с клеткой, но предотвращают слияние клеток и их инфицирование.
В исследованиях ВИЧ-1 и ВИЧ-2 было обнаружено, что gp41 содержит так называемый «пептид слияния» (fusion peptide), от которого решающим образом зависит слияние клеток. Трансмембранный домен gp41 также принимает важное участие в этом процессе. При слиянии клеточных мембран могут играть роль и дополнительные области gp41, в том числе участки «цитоплазматического хвоста» молекулы.
В механизме синцитиообразования участвует структура тримерной биспирали (или пучок из шести витков), которую образуют нативные оболочечные белки вируса. Конформация gp41, которую этот белок принимает непосредственно перед процессом слияния мембран клеток не определена, но, по-видимому, она отличается от шестивиткового спирального пучка.
Таким образом, спиральная структкра в центре триммера gp41 образуется при переходе гликопротеинов оболочки ВИЧ-1 из исходного состояния в вирионе к промежуточной конформации в процессе установления связи с рецепторами на клетке.
Неясно, может ли gp41 сам по себе, с помощью домена слияния, осуществлять индукцию синцитиообразования. Кроме того, до сих пор неизвестно, какой эпитоп вирусного белка gp41 непосредственно участвует в слиянии клеток, а какой только оказывает влияние на этот патологический процесс.

Рассматривают возможное участие в слиянии клеток таких клеточных мембранных белков, как интегрины, функциональный антиген лейкоцитов-1 (LFA-1), гликопротеин CD7. Моноклональные антитела к этим поверхностным белкам клетки предотвращают агрегацию клеток и/или образование синцития. Но при этом антитела против LFA-1 не предотвращают инфицирование вирусом, в отличие от антител против CD7.
Наконец, при смешивании двух вариантов трансгенных клеток мыши, одни из которых экспрессируют человеческую молекулу CD4, а вторые инфицированы ВИЧ, образования синцития не происходит. Эти эксперименты указывают на то, что, по-видимому, для образования синцития недостаточно взаимодействия молекулы CD4 с оболочечными белками вируса, необходим(ы) еще какой-то (какие-то) клеточный фактор(ы), имеющийся в клетках человека, но отсутствующий в клетках мыши.
В одном из исследований было установлено, что этим фактором, предположительно, является некий гликолипид. Эти данные поддерживают представление, что молекулярные механизмы процессов слияния клеток и слияния вируса с клеткой не идентичны.
Процесс слияния клеточных мембран связывают с цитопатогенными свойствами вируса и гибелью клеток. Некоторые исследователи считают, что слияние клеток приводит к уменьшению количества CD4+-клеток вследствие лизиса отдельных клеток и образования многоядерных клеток.
Механизмы гибели могут быть связаны с нарушением целостности клеточных мембран. Многоядерные клетки in vivo не обнаружены ни в каких тканях инфицированных людей, за исключением мозга. В случае других ретровирусов, многоядерные клетки могут сохранять жизнеспособность в течение длительного периода.
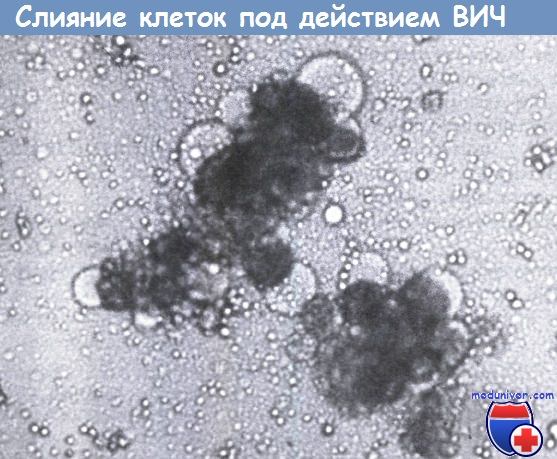
Также можно видеть пузырчатую дистрофию клеток. Фа-зово-контрастная микроскопия; увеличение, х80.
- Читать "Накопление вирусной ДНК ВИЧ как причина смерти клетки"
Оглавление темы "Инфицирование ВИЧ":- Инфицирование ВИЧ покоящихся CD4-лимфоцитов
- Инфицирование ВИЧ покоящихся CD4-эпителиальных клеток
- Клеточная латентность ВИЧ-инфекции и ее активация
- Механизмы развития латентной ВИЧ-инфекции
- Клинически латентное состояние ВИЧ
- Основные моменты теории репликации ВИЧ в клетках
- Цитопатогенные свойства ВИЧ
- Слияние клеток под действием ВИЧ - образование синцития
- Механизм слияния клеток в синцитий при ВИЧ
- Накопление вирусной ДНК ВИЧ как причина смерти клетки