Строение вируса (вириона) ВИЧ
При рассмотрении под электронным микроскопом ВИЧ-1 и ВИЧ-2 обнаруживают характерные черты лентивирусов с конусообразной сердцевиной (нуклеоид или капсид), оболочка которой состоит из белка с мол. массой 24 кДа - р24, принадлежащего группо-специфичному подмножеству вирусных белков (Gag - groop-specific antigen).
Белок оболочки капсида - р24 обозначают еще как «СА». По традиции вирусные белки обозначают буквами «р» (от protein) или gp (от glycoprotein) с цифрой, соответствующей молекулярному весу белка в килодальтонах (или в относительных единицах, х 1000). Диаметр вириона составляет от 100 до 120 нм, а его морфологические формы гетерогенны.
Оболочка вирионов в основе является мембраной человеческих клеток, в которых происходила репликация вируса. При отпочковывании вирионов в мембрану включаются два структурных вирусных белка, называемые белками оболочки - gp 120 (самый поверхностный) и gp41 (трансмембранный). Внутри вириона располагаются три структурных белка группы Gag: матриксный (МА, р17 - расположен между оболочкой вириона и нуклеоидом), р24 (СА - составляет оболочку нуклеоида) и нуклеокапсидный белок р7 (NC).
Р17 образует внутреннюю оболочку частицы, расположенную непосредственно под вирусной мембраной, СА образует конусообразную сердцевину, покрывающую РНК вирусного генома, а NC связан с вирусной РНК внутри капсида. Эти вирусные белки образуются с помощью вирусной протеазы (PR), которая расщепляет полипептидный предшественник Gag-белка р55 ВИЧ-1. Gag-белки располагаются в молекуле белка-предшественника р55 в следующем порядке: р17-р24-р2-p7-p1-p6.
Нерасщепленный р55 Gag содержит три основных домена, которые обозначают как мембрансвязывающий (М), взаимодействующий (I) и поздний (L). Домен М, локализованный в области МА, миристилирован и отвечает за прикрепление большого транслята Gag-белка к плазматической мембране клетки-хозяина. Домен I отвечает за взаимодействие мономеров Gag и располагается внутри NC области. Домен L, также локализованный в области NC, вероятно участвует в отпочковании дочерних вирионов из инфицированной клетки.
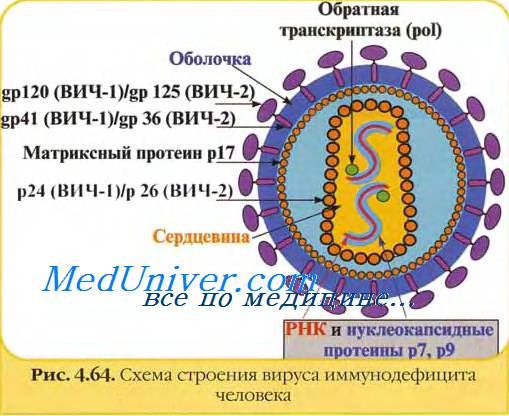
Внутри капсида, или нуклеоида, находятся две, обычно идентичные, нити РНК, с которой тесно связаны вирусная РНК-зависимая ДНК-полимераза, также носящая название обратной транскриптазы - RT, которая состоит из двух полипептидных цепей - р66 и р51. Кроме того, с геномной РНК вируса связаны NC-белки (р9 и р6). Как уже отмечали выше, внутренняя часть оболочки вируса окружена миристилированным белком р17 (МА), который является частью вирусной частицы.
Сборка ВИЧ, по-видимому, начинается с белок-нуклеинового взаимодействия в центре частицы. Матриксный белок р 17 необходим для встраивания вирусных оболо-чечных белков gp120 и gp41 в оболочку дочерних вирионов.
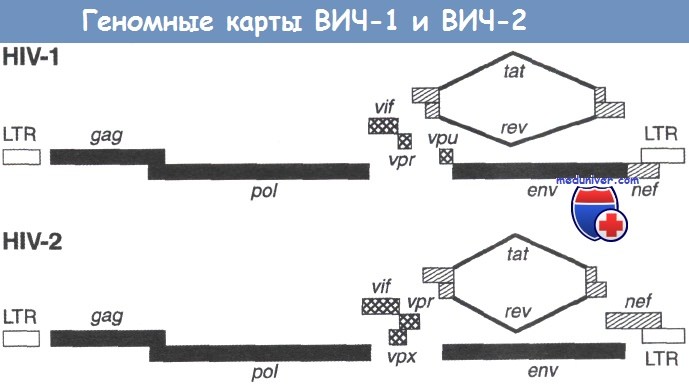
С нуклеоидом вириона частицы тесно связаны регуляторные вирусные белки Vif и Nef. На один вирион приходится от 7 до 20 молекул Vif. Также внутри вириона обнаруживают продукт добавочного вирусного гена Vpr (в ВИЧ-1) (или Vpx в ВИЧ-2), по-видимому, локализованный вне нуклеоида.
Наличие всех этих трех белков в частицах вириона говорит о том, что они играют роль на ранних стадиях инфицирования вирусом клеток. Определенные белки цитоскелета (например, актин, эзрин, эмерин, моезин и кофлин), возможно, расщепляемые вирусной протеазой, также обнаруживают в свободных вирионах.
Если эти клеточные белки и играют какую-то роль в развитии ВИЧ-инфекции, то эта роль неизвестна. Тем не менее эмерин, по-видимому, необходим для репликации ВИЧ, поскольку он обеспечивает взаимодействие и последующую интеграцию ДНК провируса в геном инфицированной клетки. Эмерин обеспечивает связь между внутренней оболочкой ядра клеток и хроматином. Кроме того, белок теплового шока hsp70, который также обнаружили внутри оболочки сердцевины ВИЧ-1, помогает поддерживать структурную целостность нуклеоида.
Наконец, как и у других ретровирусов, в вирионах ВИЧ обнаруживают селективные включения липидных доменов из мембраны клеток хозяина, которые встраиваются при отпочковании вируса. Эти включения отражаются в молярном соотношении холестерола и фосфолипидов.
Вирусные белки оболочки (Env - Envelope) образуются из гликопротеидного предшественника с мол. массой 160 кДа, gp160, который расщепляется внутри клетки (скорее всего, с помощью клеточных ферментов в аппарате Гольджи) на gp120, белк внешней поверхности оболочки вируса и трансмембранный белок gp41.
Первоначальные описания структуры поверхности вируса указывали на наличие до 72 «узелков», содержащих эти белки оболочки (Env) в виде триплетов. Однако к моменту высвобождения вируса из клетки на его поверхности обнаруживают уже всего лишь от 7 до 14 триплетов gp120/gp41 (шипов Env), которые видны на поверхности вируса при электронно-микроскопическом исследовании и при проведении криоэлектронной микроскопической томографии. Эти «шипы» организованы в структуры, подобные треножникам, на поверхности вируса.
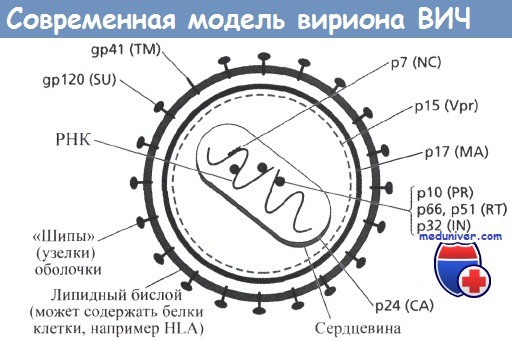
Точная локализация белков Nef и Vif, связанных с сердцевиной, до сих пор не установлена.
Использованы рекомендованные сокращения названий вирусных белков.
Небольшое число белков оболочки на поверхности вириона скорее всего отражает снижение синтеза Env в клетке, а не более позднее сбрасывание сбрасывание оболочечных белков. Атомная силовая микроскопия, однако, показывает, что белки оболочки находятся на поверхности вириона в количестве от 70 до 100 пучков, каждый из которых представляет собой тример gp41 в соединении с различным числом мономеров gp120, которые не находятся в классической тримерной форме. Таким образом, настоящее строение оболочки инфекционного вириона остается невыясненным.
Белки оболочки транспортируются на поверхность клетки, где часть центральной и аминоконцевой последовательности gp41 экспрессируется на внешнюю сторону будущего вириона. Петлевой сегмент gp41 необходим для слияния с мембраной. Центральная область этого трансмембранного белка нековалентно связывается с внешним вирусным gp120, преимущественно с его двумя гидрофобными областями, расположенными на аминоконцевом и карбоксиконцевом участках молекулы gp120. Длинный цитоплазматический «хвост» gp41, по-видимому, необходим для встраивания гликопротеина оболочки ВИЧ-1 в вирионы.
Результаты некоторых исследований позволяют предположить, что узелково-ячеистая структура необходима вирусу и некоторые области оболочки, в частности V1/V2, С2 и V3 домены gp120, помогают стабилизировать образующуюся связь. Недавно было показано, что так называемый «с хвостом взаимодействующий белок» с мол. массой 47 кДа (TIP47 - tail-interacting protein 47) связывается как с вирусными оболочечными белками, так и с матриксным белком р17, образуя четвертичную структуру белкового комплекса, обеспечивающую правильное встраивание вирусных оболочечных белков в отпочковывающийся вирион.
Белок gp120, локализованный на внешней поверхности вируса, содержит область (области) связывания клеточного рецептора (клеточных рецепторов) и главные домены для нейтрализующих антител. Наружная часть gp41 также обладает чувствительностью к нейтрализующим антителам. Обычно вирион содержит в 60-100 раз больше белка оболочки нуклеоида р24, чем белка внешней вирусной оболочки gpl20 и в 10-20 раз больше молекул р24, чем молекул полимеразы.
Во время этого процесса может происходить встраивание поверхностных белков клетки в поверхностный слой вируса, и, по-видимому, это является одной из причин появления ложноположительных реакций при проведении твердофазного иммуноферментного анализа.
Антитела к нормальным Т-клеточным белкам (например, HLA и CD4) могут давать положительную реакцию. Микрофотография любезно предоставлена Н. Gelderblom.
Микрофотография предоставлена R. Munn.

c Не обнаружен внутри вириона
d Обнаружен только в ВИЧ-1
e Закодирован только в ВИЧ-2. Возможно, является двойником Vpr
*Есть и другое мнение: NC (р7 или р9) выполняет роль шаперона для геномной РНК ВИЧ и защищает её от разрушения РНКазой Н.
**Rev выводит вирусную РНК несплайсированной из ядра, что дает возможность части РНК стать геномной для дочерних вирионов.
***Nef защищает зараженные клетки от преждевременного апоптоза; ингибирует на клетках экспрессию MHC-I, чем защищает инфицированные клетки от атаки цитотоксических Т-лимфоцитов.
****Vpr входит в состав преинтеграционного комплекса и обеспечивает продвижение этого комплекса в ядро
- Вернуться в оглавление раздела "Инфекционные болезни"
Оглавление темы "ВИЧ":