Биологические свойства недавно переданного ВИЧ
Филогенетический и молекулярный анализ общих (идентификационных) последовательностей нуклеотидов в генах, кодирующих белки вирусной оболочки (особенно V3-петли) используется для демонстрации недавно произошедшей передачи одного изолята ВИЧ-1 другому человеку и от матери к ребенку.
В большинстве случаев была показана селективная передача одного или ограниченного количества вариантов ВИЧ-1. Напротив, у реципиентов одного препарата факторов свертывания данные секвенирования ПЦР-продуктов показали отсутствие существенных различий между вирусами, изолированными от разных пациентов.
Этот метод также помог доказать инфицирование пяти пациентов по вине стоматолога во время проведения лечебных процедур. Таким образом, «входные ворота» инфекции могут существенно влиять на разнообразие вируса в организме зараженного.
Основной вопрос, который стоит перед научным сообществом, заключается в том, почему после острого и первичного инфицирования преимущественно в организме выявляются R-5-вирусы. Некоторые исследователи считают, что преимущественное распространение этих вирусов связано с их способностью инфицировать дендритные клетки слизистых оболочек или с присутствием больших концентраций SDF-1 на слизистых оболочках.
Эти объяснения не дают удовлетворительного ответа на поставленный вопрос, поскольку при первичной трансмиссии обнаруживаются оба фенотипа вируса. Х4-вирусы также могут инфицировать дендритные клетки, а R5-вирусы появляются преимущественно при первичном инфицировании путем внутривенного введения наркотиков, в случае которого основной мишенью, скорее всего, являются CD4+-клетки. Способность к распространению ВИЧ, по-видимому, не определяет результат; скорее окружающая среда влияет на то, какой тип вируса обнаруживается после острого инфицирования. Например, активация иммунной системы приводит к усилению экспрессии CCR5 и может способствовать персистенции R5-вируса.
Предпочтительное появление определенного фенотипа вируса на ранней стадии инфекции, по-видимому, не зависит от общего уровня экспрессии CCR5 и CXCR4 в CD4+-клетках периферической крови во время первичной передачи инфекции. Практически все циркулирующие наивные CD4+-клетки экспрессируют CXCR4, а не CCR5 и, таким образом, более чувствительны к Х4-вирусам. Активированные Т-клетки памяти экспрессируют CCR5. Таким образом, активация иммунитета, вызванная острым инфицированием, может создавать благоприятные условия для появления R5-виpyca.
Кроме того, может играть роль высокая активность продукции дочерних вирусных частиц R5 вирусов в МКПК и лимфоидных тканях. В культуре клеток, однако, репликация Х4-вирусов может происходить быстрее, при этом образуется большее количество дочерних вирусных частиц за единицу времени, чем при репликации R5-вирусов. Кроме того, R5-вирусы могут реплицироваться в менее активных CD4+-клетках. В связи с этим ингибиторы дополнительной стимуляции вызывают прекращение репликации Х4-, но не R5-вирусов, наиболее вероятно, за счет уменьшения импорта преинтеграционного комплекса вируса в ядро. Эти данные говорят в поддержку того, что R5-вирусы легче инфицируют слабо активированные клетки.
Кроме того, CCR5 экспрессируется на активированных CD4+-Т-клетках после взаимодействия с дендритными клетками. Эффективное распространение R5-вирусов от дендритных клеток на Т-клетки также может происходить с помощью конъюгатов, образующихся при взаимодействии вируса с дендритной клеткой.
Таким образом, в связи с общим состоянием иммунной активации R5-вирусы во время острой инфекции имеют достаточное количество чувствительных мишеней для инфицирования и репликации.
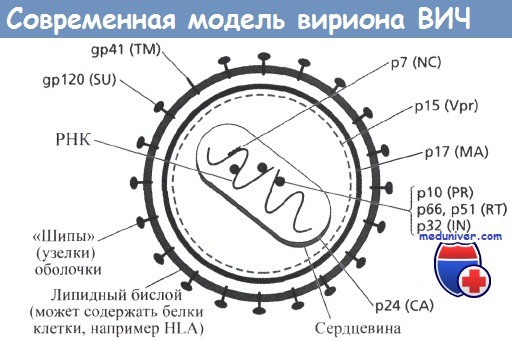
Точная локализация белков Nef и Vif, связанных с сердцевиной, до сих пор не установлена.
Использованы рекомендованные сокращения названий вирусных белков.
Важно отметить, что R5-вирусы, по-видимому, инфицируют преимущественно CCR5-экспрессирующие CD4+-клетки ЖКТ, который представляет собой обширный резервуар этих клеток. Кроме того, R5-вирусы могут преимущественно передаваться на CCR5-CD4+-клетки с клеток слизистой оболочки кишечника. Выраженная вирусемия, наблюдающаяся при острой инфекции, скорее всего, связана с таким тканевым инфицированием и гибелью CD4+-клеток. Исследования на макаках резус показали, что R5 SHIV в большей степени, чем Х4 SHIV, вызывает гибель CD4+-Т-клеток кишечника и позднее происходит уменьшение количества периферических CD4+-клеток.
Напротив, Х4 SHIV-инфекция поражает преимущественно периферические CD4+-клетки и клетки лимфоидных тканей, но слабо поражает клетки кишечника Такие возможные различия в патогенезе R5 и Х4 вирусных инфекций могут быть связаны с влиянием иммунной системы организма-хозяина.
R5-виpyc, возможно, слабее распознается иммунной системой (например, ЦТЛ), в основном за счет того, что он инфицирует макрофаги и дендритные клетки. Это предположение возникло при проведении исследований, в которых уничтожали CD8+-клетки у макак резус, совместно инфицированных R5- и Х4-вирусами. При этом появлялся Х4-вирус, что указывает на дифференциальную борьбу CD8+-Т-клеток с этим вирусом.
Два примечательных случая случайной передачи вируса позволили несколько прояснить этот вопрос. В обоих передача происходила не через слизистые оболочки. В одном из них передача произошла в результате внутривенного введения небольшого количества крови, содержащей преимущественно Х4-вирус. В другом было внутримышечно введено большое количество крови, содержащей преимущественно вирус R5. В первом случае развилась быстрая устойчивая репликация Х4-вируса, которая сопровождалась быстрой гибелью CD4+-Т-клеток. Во втором - передача преимущественно R5-вируса первоначально привела к селективному увеличению численности Х4-вируса до сероконверсии.
Затем, после сероконверсии, появились R5-вирусы и количество CD4+-T-клеток уменьшилось. Преимущественное замещение Х4-вируса 115-вирусом также наблюдалось другими исследователями и в одном случае было связано с присутствием нейтрализующих антител в организме хозяина, действующих на Х4-, но не на R5-вирусы. Несмотря на то, что феномен стенотипического переключения при первичном изолировании встречается редко, он показывает, каким образом организм хозяина через клетки-мишени или иммунные реакции может влиять на ранние стадии инфицирования вирусом и на его распространение.
Наконец, существуют некоторые доказательства того, что секретируемый вирусный белок Tat может ингибировать Х4-вирусы через взаимодействие с CXCR4 и таким образом способствовать преобладанию R5-вирусов. Эти данные отличаются от данных об активации обоих типов хемокиновых корецепторов под действием Tat, хотя, нужно отметить, что он может стимулировать выработку других клеточных факторов, в частности SDF-1, который способен блокировать экспрессию CXCR4.
Причины преобладания R5-вируса в период острой и первичной инфекции:
• Для R5-вирусов во время иммунной активации существует большое количество чувствительных CCR5+-CD4+-Т-клеток мишеней
• Активация иммунной системы усиливает экспрессию CCR5
• R5-вирусы могут эффективно инфицировать неактивированные клетки
• R5-вирусы могут инфицировать макрофаги и дендритные клетки
• Высокая концентрация R5 дочерних вирусных частиц в инфицированных клетках
• R5-вирусы преимущественно инфицируют CCR5-экспрессирующие CD4+-клетки в желудочно-кишечном тракте
• R5-вирусы могут передаваться от экспрессирующих CCR5-клеток слизистой оболочки кишечника на CD4+-Т-клетки
• R5-вирусы слабее распознаются иммунной системой (например, ЦТЛ)
• Секретируемый вирусный белок Tat может ингибировать Х4-вирусы, взаимодействуя с CXCR4
• Преинтеграционный комплекс R5-вирусов может легче встраиваться в ядро неактивированных клеток
• Взаимодействие CD4+-клеток с дендритными клетками усиливает экспрессию CCR5
- Читать "Переключение фенотипа ВИЧ с R5 на X4 в процессе развития инфекции"
Оглавление темы "Течение и прогноз ВИЧ":- Влияние генетических факторов на длительность жизни при ВИЧ
- Влияние типа вируса на длительность жизни при ВИЧ
- Влияние иммунного ответа на длительность жизни при ВИЧ
- Варианты клинического течения SIV-инфекции (вирусного иммунодефицита обезьян)
- Особенности людей устойчивых к ВИЧ (сероотрицательные лица с высоким риском заражения)
- Генетические особенности людей устойчивых к ВИЧ
- Клеточный иммунный ответ людей устойчивых к ВИЧ
- Гуморальный иммунный ответ людей устойчивых к ВИЧ
- Биологические свойства недавно переданного ВИЧ
- Переключение фенотипа ВИЧ с R5 на X4 в процессе развития инфекции