Белок TRIM5a как механизм резистентности к ВИЧ
В нескольких ранних исследованиях было показано наличие каких-то противовирусных механизмов в клетках приматов, которые способны 50-кратно ограничить репликацию вируса лейкемии мышей, тропного к клеткам перевивной линии NIH (N-MLV), в клетках человека и репликации ВИЧ в клетках макаки резус. В клетках человека ген, отвечающий за блокировку репликации N-MLV, а также вируса инфекционной анемии лошадей (EIAV), получил название Ref-1 (resistance factor-1 - фактор устойчивости-1).
По-видимому, Ref-1 действует на стадии, предшествующей обратной транскрипции. В клетках обезьян ген, получивший название Lv-1 (сокращенно от лентивирусного фактора чувствительности 1 - lentivirus susceptibility factor 1) считается ответственным за блокировку синтеза ДНК ВИЧ до начала обратной транскрипции.
Как ограничение репликации MLV в клетках человека под действием Ref-1, так и ограничение репликации ВИЧ в клетках обезьяны под действием Lv-1, по-видимому, происходят с участием белков вирусного капсида, и оба эти ограничения преодолеваются большим количеством вируса.
Различия между этими противовирусными внутриклеточными механизмами можно оценить путем тестирования разных линий клеток и использования различных вирусов. Торможение вирусной репликации у приматов напоминает функционирование гена Fv-1 у мышей, который определяет тропизм различных типов вирусов лейкемии мышей (MLVs). Fv-1 кодирует клеточный белок, который взаимодействует с капсидным белком вирусов и тем самым предотвращает репликацию вируса до начала обратной транкрипции. По-видимому, ген Fv-1 сам имеет ретровирусное происхождение.
Объяснение дополнительных механизмов внутриклеточного подавления репликации ретровирусов у приматов было найдено после того, как было показано, что в качестве действующего внутриклеточного фактора выступает цитоплазматический белок TRIM5a (tripartite motif protein 5a). Белки семейства TRIM состоят из N-концевого домена RING, одного или двух В-бокс-доменов (В box) 1 или 2 типа и сильно спирализованного участка.
Было показано, что гены TRIM5a контролируют Ref-1 -зависимое ограничение репликации MLV и EIAV в клетках человека и Lv-1 -зависимое торможение репликации ВИЧ в клетках обезьян. Примечательно, что TRIM5a клеток макаки резус ограничивает репликацию ВИЧ-1, но не SIVmac.
TRIM5a в клетках человека не останавливает репликацию ВИЧ или других ретровирусов приматов. Кроме того, было обнаружено, что в клетках африканской зеленой обезьяны происходит торможение репликации ВИЧ-1, ВИЧ-2, SIVmac, N-MLV и EIAV, но не SIVagm. Было показано, что и в этом случае наблюдаемый эффект связан с продуктом гена TRIM5a. Такая природная противовирусная защита клеток является видоспецифичной (как и в случае APOBEC3G) и, скорее всего, связана с торможением высвобождения сердцевины вируса из оболочки.
В этом отношении она напоминает действие Fv-1. Другой возможный механизм действия TRIM5a может заключаться в наведении на вирусные частицы клеточных механизмов деградации). TRIM5a действует на более ранней стадии обратной транскрипции, чем APOBEC3G, и не встраивается в вирионы.
Ограничение репликации ВИЧ в клетках приматов связано с различиями во взаимодействии вирусного капсида с СурА.
Этот собственный белок клетки связывается с белком нуклеокапсида ВИЧ-1 и способствует развитию инфекции в случае многих изолятов ВИЧ-1 (например, группы М). Обработка клеток человека и обезьяны циклоспорином, антагонистом СурА, приводит к различным последствиям.

В клетках обезьяны циклоспорин предотвращает взаимодействие нуклеокапсида вируса с СурА и может более чем в 100 раз усилить репликацию ВИЧ в клетках этих приматов. Поэтому такое связывание предотвращает эффект Lv-1. Напротив, в клетках человека циклоспорин блокирует репликацию ВИЧ, скорее всего, также путем разрушения комплекса р24 с СурА.
Однако в этом случае комплекс «р24-СурА», по-видимому, играет важную роль в предотвращении действия фактора, ограничивающего активность TRIM5a (Ref-1) в клетках человека. Результаты исследований с циклоспорином говорят о том, что ВИЧ использует один клеточный белок для того, чтобы противостоять другому клеточному белку, экспрессируемому в клетках человека и обладающему противовирусными свойствами.
Эти данные свидетельствуют о том, что в инфицированной клетке существует динамическое взаимодействие между белком вирусного капсида р24 и клеточными белками СурА и TRIM5a. До сих пор неизвестно, каким образом циклофилин в клетках макаки резус осуществляет торможение инфекционного процесса ВИЧ-1, но, возможно, это происходит за счет облегчения взаимодействия нуклеоида вируса с TRIM5a макаки резус.
В связи с этим было показано, что у TRIM5a, экспрессируемого у обезьян дурукули (обыкновенной ночной обезьяны), есть fusion-CypA-белок, который способствует резистентности этих клеток к инфекции ВИЧ-1.
Аналогично данным, показывающим, что замена одной аминокислоты в белке APOBEC3G приводит к изменению его функций, замена одной аминокислоты в SPRY-домене TRIM5a человека может сделать его сходным с гомологичным белком клеток приматов и привнести свойство противовирусной активности в отношении ВИЧ-1 в клетках человека.
Внутриклеточные противовирусные механизмы работают и против ВИЧ-2; инфекция этим вирусом блокируется на этапе после обратной транскрипции и до проникновения в ядро. Этот противовирусный блок определяется геном Lv-2 и его продуктом, во взаимодействии с которым участвуют вирусные белки нуклеокапсида и оболочки.

Недавние исследования позволили предположить, что этот механизм ингибиции инфекции после проникновения вещества вируса в клетку связан с рН-независимыми процессами формирования эндосом в липидных рафтах мембраны клетки, в которых со стороны вируса основным определяющим фактором являются белки оболочки вируса.
Помимо TRIM5a, другие белки семейства TRIM влияют на различные клеточные процессы, в том числе на пролиферацию клетки, дифференцировку, развитие и онкогенез. По-видимому, в геноме человека около 70 генов кодируют белки семейства TRIM. Только несколько из них охарактеризованы в достаточной мере.
Известно, что один из белков семейства TRIM, TRIM32, связывается с активационным доменом белка Tat, но функциональные последствия этого взаимодействия неизвестны. Другой член семейства TRIM, TRIM22, представляет собой индуцируемый интерфероном фактор и может подавлять экспрессию LTR ВИЧ. Его влияние на репликацию ВИЧ не установлено.
Исследования чувствительности клеток животных разных видов к инфицированию ВИЧ показали, что торможение репликации вируса в мышиных клетках происходит на стадии сборки вириона, а не на стадии обратной транскрипции. Это может быть связано с другим видом противовирусной активности. Кроме того, был обнаружен белок Murr-1, который способен обеспечивать латентное состояние вирусной инфекции за счет препятствия деградации IkB.
К тому же был открыт некий нутриклеточный фактор, ингибирующий сборку вирионов ВИЧ. По-видимому, вирусный белок Vpu инактивирует этот фактор. Исследования внутриклеточных механизмов подавления вирусных инфекций, в том числе ВИЧ предоставляют новые возможности для разработки методов противовирусного лечения.
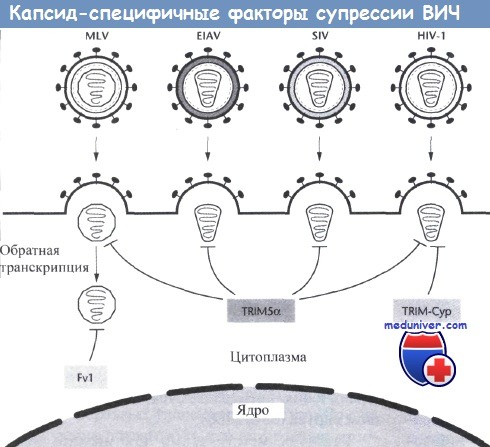
После попадания вирусных компонентов в цитоплазму клетки может происходить распознавание капсидных белков ретровирусов и дальнейшее развитие инфекции может блокироваться за счет действия одного из многих факторов.
Fv-1, уникальный мышиный белок, останавливает заражение мышиным вирусом лейкемии (MLV), и этот эффект носит исключительно Fv-1-аллель-специфичный и MLV-штаммоспецифический характер.
TRIM5a, который обнаруживается у большинства приматов, может предотвращать заражение рядом ретровирусов, в том числе N-тропным N-MLV, вирусом инфекционной анемии лошадей (EIAV), вирусом иммунодефицита обезьян (SIV) и ВИЧ-1.
Спектр противовирусной активности TRIM5a зависит от его видовой принадлежности. Уникальная форма TRIM5 появляется у обыкновенных ночных обезьян за счет транспозиции псевдогена СурА, при этом возникающий в результате fusion-белок подавляет ВИЧ-1 за счет способности последнего связываться сердцевинной частью с белком оболочки нуклеоида вируса.
- Читать "Влияние цитокинов на репликацию ВИЧ"
Оглавление темы "Репликация ВИЧ":- Этапы репликации ВИЧ внутри клетки
- Влияние внутриклеточных факторов на репликацию ВИЧ
- Особенности репликации ВИЧ в Т-клетках
- Особенности репликации ВИЧ в моноцитах и макрофагах
- Белок APOBEC3G как механизм резистентности к ВИЧ
- Аналоги белка APOBEC3G у животных
- Природные функции белков APOBEC
- Белок TRIM5a как механизм резистентности к ВИЧ
- Влияние цитокинов на репликацию ВИЧ
- Влияние белков ВИЧ на клетку