Инфицирование ВИЧ покоящихся CD4-лимфоцитов
Ранее в отдельных статьях мы обсуждали этапы развития продуктивной вирусной инфекции и ее последствия. Другой важный вопрос касается событий, происходящих при заражении ВИЧ «покоящегося» лимфоцита. Первые эксперименты с ВИЧ-инфицированием лимфоцитов показали, что репликация вируса лучше всего происходит при антиген- или митоген-индуцированной активации. Если затормозить пролиферацию клеток перевивной линии с помощью фармакологических препаратов, то в некоторых клетках репликация ВИЧ прекращается, а в других не изменяется.
Однако эти системы не отражают ситуации, которая складывается в нормальных условиях в периферических CD4+-лимфоцитах периферической крови. В некоторых исследованиях было показано, что активированные CD4+-клетки, инфицированные ВИЧ, могут переходить обратно в неактивированное состояние, на что указывают известные поверхностные маркеры клетки (например, CD69 CD25 Ki67 ).
Такие наблюдения в культурах клеток заслуживают дальнейшего изучения с целью использования в качестве модели для исследования латентного состояния ВИЧ в нативных CD4+-лимфоцитах.
1. CD4-клетки периферической крови и ВИЧ in vito
Ранние исследования с использованием неделящихся CD4+-T-клеток периферической крови позволили получить ценные сведения относительно процессов, участвующих в эффективной вирусной инфекции и продукции вирусного потомства. Результаты показали, что степень активации клетки (судя по экспрессии HLA-DR и CD25) может влиять на интенсивность обратной транскрипции с вирусного генома и на высвобождение новых вирионов из клетки.
CD4+-клетки с полностью заблокированным синтезом ДНК, находящиеся в неактивированном состоянии (в стадии G0/G1) (т.е. не экспрессирующие HLA-DR и экспрессирующие мало CD25), не подвергаются продуктивному ВИЧ-инфицированию. Несмотря на экспрессию CD4, в этих клетках не проявляются признаки вирусной инфекции и при их активации в условиях, достаточных для поддержания эффективной продукции вируса в культурах клеток in vitro.
Молекулярные исследования показали, что вирус проникает в покоющуюся клетку, но затем обычно образуются только короткие транскрипты вирусной ДНК; обратная транскрипция осуществляется медленно или не полностью и не может активироваться после проникновения вируса. Преинтеграционные комплексы находятся в лабильном состоянии. Таким образом, инфекция носит абортивный характер, и процессы катаболизма в клетке снижают репликативную способность инфекционного вируса в течение от 1 до 2 дней.
Некоторые исследователи показали, что покоящиеся CD4+-лимфоциты могут инфицироваться in vitro, при этом репликация вируса сначала подавлена, но может быть активирована. Определение «покоящиеся» имеет важное значение, поскольку эти клетки могли ранее находится в активном состоянии и в них могут сохраняться внутриклеточные факторы, экспрессия которых необходима для развития ВИЧ-инфекции. Неинтегрированные формы провирусной ДНК можно определить с помощью ПЦР в таких покоящихся Т-клетках в период от нескольких дней до двух недель после экспозиции с вирусом.
После активации эти клетки начинают выделять инфекционно дееспособные дочерние вирусы. Данные исследования позволили предположить, что вирус проникает в «покоящуюся» клетку, но транскрипция вирусных генов носит ограниченный характер. Провирусная ДНК не интегрируется в геном клетки-хозяина, и вирусные белки не вырабатываются. В определенных случаях, если эти клетки не активируются в течение трех-пяти дней, может произойти купирование инфекции. В других ситуациях в клетке могут производиться длинные формы провирусной ДНК, инфекция принимает непродуктивный характер и способна персистировать от двух до четырех недель. При таком типе инфицирования преинтеграционные комплексы остаются в цитоплазме до тех пор, пока активация не приведет к быстрой транспортировке PIC в ядро, процесс которой требует АТФ и не требует деления клетки. После активации образуется полноразмерная провирусная ДНК, которая эффективно интегрируется в хромосомы, после чего начинается продукция новых вирусных частиц.
Сравнение состояний покоя и активации клетки в отношении поддержания вирусной инфекции представляет существенный интерес. Покоящиеся клетки, не экспрессирующие CD25, проявляют невосприимчивость к ВИЧ-инфекции, наиболее вероятно на стадии проникновения вируса в клетку, поскольку в клетках не удается обнаружить никаких следов вирусных транскриптов. Однако при совместном культивировании с CD257 CD4+-клетками все-таки происходит непродуктивное инфицирование покоящихся CD25 -клеток, сходное с описанным выше.
Некоторые факторы, вырабатываемые CD25+-клетками (например, IL-2) могут оказывать влияние на этот процесс, увеличивая чувствительность клеток-мишеней к инфекции.
Другие эксперименты показывают, что ВИЧ способен проникать в выделенные CD45RA+ (покоящиеся лимфоциты) или CD45RO+ (лимфоциты памяти) CD4+-клетки, но только в CD45RCT CD4-клетках (лимфоцитах памяти) со временем наблюдается продуцирование вируса. Эти исследования позволили предположить, что вирус проникает в неиммунные CD45RA+ клетки, но до тех пор, пока на клетку не подействуют стимулы, вызывающие ее активацию, инфекция протекает в абортивной форме, что, скорее всего, связано с подавлением транспортировки преинтеграционных комплексов вируса в ядро. Эти результаты не согласуются с данными, которые показывают, что экспрессия хемокиновых рецепторов на неиммунных (CXCR4) клетках, в отличие от клеток памяти (CCR5+) может влиять на вирусную инфекцию.
В упомянутых исследованиях оба типа клеток экспрессировали CXCR4 и проявляли чувствительность к вирусу биотипа Х4. Однако состояние активации определяет степень репликации ВИЧ, как было показано в других работах, описанных выше. Эти данные подтверждают и помогают объяснить результаты предыдущих исследований, которые говорят о предпочтительном инфицировании ВИЧ-1 CD4+-клеток памяти.
В других экспериментах, проведенных на покоящихся CD4-клетках, было показано, что ВИЧ-инфицирование макрофагов приводит к высвобождению факторов, которые взаимодействуют с В-клетками, вызывая секрецию IL-6, что делает покоящиеся CD4+-клетки пригодными для поддержания репликации вируса. Эти данные требуют дальнейшего подтверждения, но на их основании уже можно предположить возможные механизмы инфицирования покоящихся клеток; такие клетки способны играть роль длительно существующих резервуаров ВИЧ в организме.
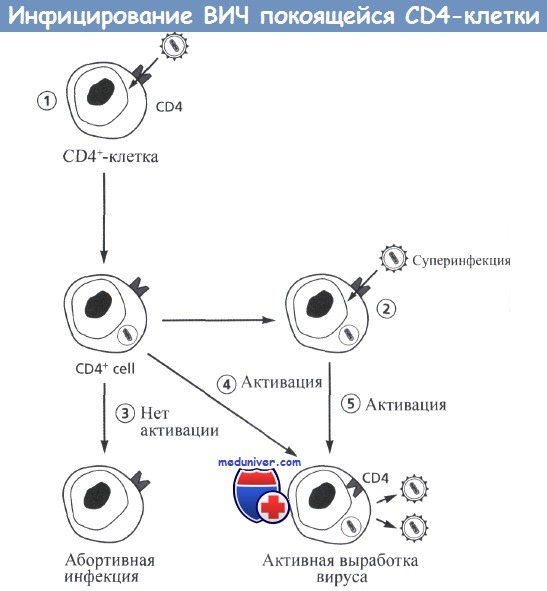
Если такая клетка не активируется, репликации вируса не происходит. Поскольку молекула CD4 продолжает экспрессироваться в покоящейся клетке, может произойти суперинфицирование другим вирусом (этап 2).
Если после инфицирования не произошло активации покоящейся клетки, развивается абортивная форма инфекции (этап 3).
При активации клетки начинается продуктивная репликация вируса (этап 4).
Сходным образом активация подвергшейся суперинфицированию клетки может привести к продукции вируса, при этом могут обнаруживаться вирусы обоих типов и рекомбинантные формы.
После активации и продукции вируса (этап 5) происходит угнетение экспрессии молекулы CD4 и подавление развития суперинфекции.
2. Покоящиеся CD4-клетки и ВИЧ in vivo
Данные, полученные in vitro при изучении очищенных покоящихся CD4+-клеток, также были проверены в клинических исследованиях. Очищенные CD4+-клетки, полученные от людей на бессимптомной стадии инфекции, в основном состояли из покоящихся клеток, содержащих преимущественно неинтегрированную провирусную ДНК. На большинстве этих клеток не обнаруживали экспрессию HLA-DR, но на небольшом проценте клеток (0,3%) молекулы HLA-DR экспрессированы. Фенотипический анализ латентно инфицированных клеток показал практически полное отсутствие признаков вирусной инфекции.
Таким образом, такие клетки плохо распознаются иммунной системой. Если пациент получает высокоактивную противоретровирусную терапию, количество латентно инфицированных CD4+-клеток может уменьшаться до 1 миллиона на 10 миллионов клеток; после прекращения лечения реплицирующийся вирус будет обнаруживаться лишь в небольшой доле клеток. Такие латентно инфицированные клетки появляются на ранних стадиях острой ВИЧ-инфекции даже при проведении лечения.
Данные, полученные при изучении покоящихся клеток, говорят в пользу того, что у большинства пациентов на бессимптомной стадии заболевания ВИЧ присутствует в неинтегрированной неинфекционной форме после проникновения в клетку, возможно, в течение гораздо более длительного времени, чем было показано в исследованиях на культуре клеток in vitro. Другие клетки крови? также могут содержать интегрированную, но не экспрессирующуюся вирусную мРНК или даже вирусные белки. Возможно, подавление вирусной репликации в покоящихся клетках in vivo связано с действием CD8+-клеток, вырабатывающих анти-ВИЧ фактор, подавляющий репликацию вируса, и, возможно, что этот механизм вносит существенный вклад в «арест» репликации вируса в неактивированных клетках in vivo. Предположительно свободные вирионы в плазме крови являются источником повторных заражений покоящихся клеток, которые элиминируют вирусные нуклеиновые кислоты до тех пор, пока не будут чем-либо активированы.
Если репликации вируса не происходит, степень экспрессии CD4 на клетках может сохраняться на прежнем уровне, в связи с чем возможно повторное инфицирование или суперинфицирование. Таким образом, циклические флуктуации вирусной инфекции могут характеризовать динамическое состояние, наблюдающееся в организме инфицированного человека. В таких условиях существует вероятность рекомбинации разных вариантов вирусов при множественном инфицировании клетки.
3. Внутриклеточный механизм инфицирования CD4-клеток ВИЧ
Были идентифицированы внутриклеточные факторы, которые могут влиять на степень продукции вируса в покоящихся CD4+-клетках. Ими оказались белки клетки, которые связываются с ДНК (например, NF-kB) и/или РНК, которые могут тормозить репликацию ВИЧ.
Фактор транскрипции NF-kB, как правило, рассматривают в качестве существенного участника про-воспалительных процессов, в результате которых более ожидаемым является усиление репликации вирусов. Вероятно, такие параметры не подлежат однозначному толкованию.
Кроме того, как отмечали выше, результаты, полученные с R5- и Х4-изолятами ВИЧ-1, говорят о блокировке обратной транскрипции в активированных Т-клеточных линиях и макрофагах соответственно, что говорит о возможном участии сходных внутриклеточных механизмов.
Некоторые наблюдения, сделанные in vitro и in vivo, касающиеся ограничения обратной транскрипции ВИЧ в покоящихся клетках, могут объясняться по-другому. Например, в вирионах ВИЧ иногда можно обнаружить различные фрагменты молекул провирусной ДНК, что указывает на существование явления неполноценной обратной транскрипции. Наличие такой ДНК может быть причиной получения положительных результатов ПЦР-анализа, которые указывают на низкую концентрацию провирусной ДНК в покоящихся клетках. Фрагменты ДНК, полученные в результате неполноценной обратной транскрипции в клетках, могут входить в состав вирусов, связанных с клеткой, но не интернализованных, или вирусов с незавершенным циклом репликации. Однако такая возможность представляется маловероятной, поскольку в контрольных культурах, использованных для экспериментов, было обнаружено инфицирование покоящихся клеток.
Совсем недавно была показана связь торможения вирусной репликации в покоящихся CD4+-клетках с высоким уровнем экспрессии определенной разновидности APOBEC3G, а именно в покоящихся клетках обнаруживают высокое содержание низкомолекулярной формы APOBEC3G, которая тормозит репликацию вируса за счет неизвестного механизма. После активации клетки низкомолекулярная форма APOBEC3G превращается в высокомолекулярный комплекс, в форме которого APOBEC3G функционирует в нормальных условиях.
Резюме об инфицировании покоящихся CD4-клеток ВИЧ
Трудности, возникающие при объяснении данных, полученных на покоящихся клетках, связаны в первую очередь с определением «состояния покоя». Большинство иммунологов настаивает на том, что у таких клеток должна отсутствовать экспрессия CD25. Другие также подчеркивают значение отсутствия экспрессии CD69, Ki67 и HLA-DR. Если в популяции клеток in vitro присутствует небольшое количество HLA-DR+-клеток, они могут подвергаться ВИЧ-инфицированию, что может приводить к ошибочному предположению, на основании данных ПЦР-анализа, об успешном инфицировании всех клеток. Такую возможность можно исключить путем удаления всех HLA-DR+-клеток. Эти данные говорят в поддержку вывода о том, что проникновение и экспрессия вируса зависят от состояния активации клетки. Продолжительность периода времени, в течение которого клетка может нести неинтегрированную вирусную ДНК, также может коррелировать со степенью активации клетки.
Также неизвестно, каким образом покоящиеся CD4+-клетки, в которых отсутствует экспрессия HLA-DR, инфицируются in vivo. Необходимо выяснить, способны ли HLA-DR+-CD4-лимфоциты после инфицирования возвращаться в непродуктивное HLA-DR- состояние покоя. Некоторые исследования свидетельствуют о том, что такой переход возможен, но, по всей видимости, происходит крайне редко. Предположительно инфицирование может происходить при действии определенных цитокинов на покоящиеся CD4+-клетки. С другой стороны, как отмечали выше, небольшое количество покоящихся CD4+-клеток с признаками инфекции in vivo могут представлять собой небольшое количество HLA-DRf клеток, находящихся в покоящейся клеточной популяции (0,3%). Наконец необходимо выяснить клиническую значимость результатов исследований, проведенных на покоящихся Т-клетках.
Большинство CD4+-клеток in vivo находятся в состоянии относительного покоя, но при этом восприимчивы к ВИЧ и способны поддерживать продуктивную вирусную инфекцию. В связи с этим большое значение имеют факторы и события, вызывающие активацию клеток, в результате чего создаются условия, закрепляющие и интенсифицирующие вирусную инфекцию.

- Читать "Инфицирование ВИЧ покоящихся CD4-эпителиальных клеток"
Оглавление темы "Инфицирование ВИЧ":- Инфицирование ВИЧ покоящихся CD4-лимфоцитов
- Инфицирование ВИЧ покоящихся CD4-эпителиальных клеток
- Клеточная латентность ВИЧ-инфекции и ее активация
- Механизмы развития латентной ВИЧ-инфекции
- Клинически латентное состояние ВИЧ
- Основные моменты теории репликации ВИЧ в клетках
- Цитопатогенные свойства ВИЧ
- Слияние клеток под действием ВИЧ - образование синцития
- Механизм слияния клеток в синцитий при ВИЧ
- Накопление вирусной ДНК ВИЧ как причина смерти клетки