Виды биологических мембран. Разновидности клеточных липидов
Биологические мембраны не ведут себя подобно чистым липидным бислоям. Это имеет место в значительной степени в результате их холестеринового содержания, которое может составить до 50 %. Ригидные, плоские стероидные кольца взаимодействуют и иммобилизируют те области цепей жирных кислот, которые являются самыми близкими к поверхности мембраны. Это обеспечивает создание липидного бислоя с меньшим количеством жидкости и снижает его проницаемость к малым молекулам. Однако, кроме снижения жидкостных свойств, это также подавляет упаковку остатков жирных кислот и поэтому подавляет возможные фазовые переходы, что ведет к состоянию «геля».
Важно отметить, что для сохранения липидной асимметрии необходима структурная целостность цитоскелета, которая может нарушаться в клетках с дефицитом спектрина или белка полосы 4.1. Физические и химические воздействия, нарушающие архитектонику цитоскелета, изменяют и липидную асимметрию. Холестерин может быть эквимолярным с фосфолипидами и является важным регулятором жидкостных свойств мембраны. Он состоит из твердой уплощенной кольцевой части и более гибкого углеводородного хвоста, который проникает через бислой к жирным кислотам. Растительные мембраны содержат другие стеролы вместо холестерина.
Липиды мембран эритроцитов делят на 3 класса: нейтральные липиды, гликолипиды и фосфолипиды.
Нейтральные липиды, составляющие до 30% (по массе) от всех липидов, включают глицериды, холестерин и эфиры холестерина. Главной фракцией является холестерин.
Гликолипиды составляют около 10% мембранных липидов и являются гликосфинголипидами. Они содержат ненасыщенный аминоспирт сфингозин, длинноцепочечную ненасыщенную жирную кислоту и полярные углеводные группы. В зависимости от вида полярной головки гликосфинголипиды разделяют на нейтральные и кислые. Нейтральные гликосфинголипиды содержат один или несколько остатков Сахаров. Кислые гликосфинголипиды (ганглиозиды) в качестве заряженной головки несут сложные олигосахариды, включающие сиаловые кислоты.
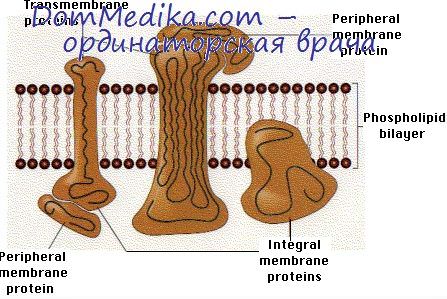
Гликолипиды ковалентно связывают углеводные цепи и сконцентрированы во внешнем слое плазматической мембраны. В животных клетках, гликолипиды образуются из церамида, предшественника сфингомиелина. Они, как полагают, играют определенную роль в клеточной адгезии и узнавании различных молекул, регуляции электрической проводимости, защитной функции и т.д.
Наиболее широко представлен в эритроцитарной мембране класс фосфолипидов (около 60%), которые могут быть производными либо сфингозина, либо глицерина. В соответствии с этим их делят на сфинголипиды, представителем которых является сфингомиелин (СМ), и глицерофосфолипиды: фосфатидилхолин (ФХ), фосфатидилэтаноламин (ФЭ), фосфатидилсерин (ФС), фосфатидилинозитол (ФИ), фосфатидная кислота (ФК). Полярные головки липидных молекул заряжены либо отрицательно, либо нейтрально; в соответствии с этим фосфолипиды разделяют на кислые (ФЭ, ФС, ФИ, ФК) и нейтральные (ФХ, СМ).
Как уже было сказано, липидный бислой асимметричен относительно липидов и белков. Асимметрия фосфолипидов обеспечивается переносчиками фосфолипидов. Гликолипиды - наиболее асимметрически распределенные липиды, потому что они только найдены на внеклеточной поверхности мембраны. Асимметричное распределение липидов функционально значимо. Так, концентрация фосфатидилсерина во внутреннем слое плазматической мембраны ведет к значительным различиям заряда между двумя поверхностями мембраны. Во внутреннем слое ФС связывает протеинкиназу С во время переноса сигнала. Фосфатидилинозитол-4,5-бисфосфат на внутренней поверхности мембраны расщепляется рецептор-активизированным ферментом до двух внутриклеточных молекул-мессенжеров, также вовлеченных в трансмембранную сигнальную передачу.
Количественное соотношение между классами липидов изменяется при некоторых заболеваниях человека. Так, например, при серповидноклеточной анемии содержание ФС, ФЭ и фосфатидной кислоты в эритроцитах несколько повышено, а содержание фосфатидилглицерина понижено. В эритроцитах больных ишемической болезнью сердца увеличивается количество холестерина и фосфолипидов, при инфаркте миокарда эти изменения менее выражены. При старении эритроцитов in vivo уменьшается содержание фосфолипидов и холестерина без изменения содержания мембранных белков.
- Читать далее "Мембранные белки. Структура мембранных белков клеток"
Оглавление темы "Структура клеточных мембран":1. Клеточные мембраны. Плазматические мембраны
2. Виды биологических мембран. Разновидности клеточных липидов
3. Мембранные белки. Структура мембранных белков клеток
4. Строение и функции белков клеточных мембран. Белки клеточных мембран эритроцитов
5. Периферические белки клеточных мембран. Спектрин клеточных мембран
6. Структура спектрина. Свойства спектрина клеточных мембран
7. Кодирование спектрина. Анкирин клеточных мембран
8. Содержание анкирина в клеточных мембранах. Анкирин в эритроцитах
9. Эритроцитарный анкириновый ген. Белок 4.1 клеточных мембран
10. Структура белка 4.1. Строение и кодирование белка 4.1