Адгезионные взаимодействия клеток друг с другом - механизмы
Адгезионные межклеточные взаимодействия играют главенствующую роль в эмбриогенезе и сохранении тканевой целостности, а также оказывают регулирующее влияние на пролиферацию клеток, их локомоцию и синтетические процессы.
Клетки образуют друг с другом контакты нескольких типов: замыкающие, или плотные, контакты (occluding, tight junctions), щелевые (gap junctions) и заякоривающие контакты (anchoring junctions).
В плотных контактах молекулы трансмембранного белка окклудина в плазматических мембранах контактирующих клеток прямо связываются друг с другом. Плотные контакты выполняют барьерные функции. При щелевых контактах между клетками остается узкая щель. Мембраны контактирующих клеток и щель между ними пронизаны цилиндрическими структурами — коннексонами (состоящими из белка коннексина), содержащими каналы, которые могут открываться и закрываться в зависимости от изменений конформации коннексина.
Благодаря каналам осуществляется метаболическая кооперация клеток — прямой межклеточный обмен ионами и небольшими молекулами без выхода их в межклеточное пространство. Заякоривающие контакты включают две разновидности: адгезионные контакты и десмосомы. По своей структуре они напоминают фокальные контакты: в их состав входят трансмембранные молекулы адгезии, которые через комплекс субмембранных белков соединяются с цитоскелетом контактирующих клеток — актиновыми микрофиламентами (в адгезионных контактах) или промежуточными филаментами (в десмосомах).
Заякоривающие контакты, в частности адгезионные (adherence junctions), играют определяющую роль в объединении клеток разных типов в тканевые структуры. Наиболее универсальными являются адгезионные контакты, в которых молекулами межклеточной адгезии служат кадхерины. Последние опосредуют адгезию клеток, принадлежащих как к одному и тому же, так и к различным типам.
Рассмотрим организацию и функции кадхеринсодержащих адгезионных контактов.
Кадхерины относятся к семейству трансмембранных гликопротеинов. Среди них кадхерины, осуществляющие адгезионные контакты эпителиальных клеток (Е-кадхерины), эндотелиоцитов (V-кадхерины) и др. За счет внеклеточных доменов своих кадхеринов клетки сцепляются друг с другом. Цитоплазматические домены кадхеринов через комплекс взаимосвязанных белков: а-, b- и у-катенинов (последние называются также плакоглобинами) , а-актинина и некоторых других белков — связываются с актиновыми микрофиламентами контактирующих клеток.
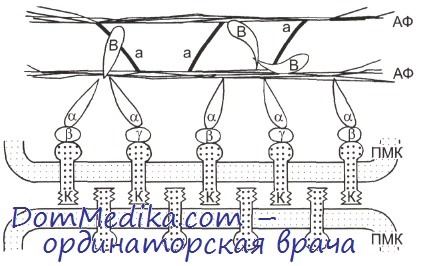
ПМК — плазматическая мембрана клетки; К — кадхерин; а, р, Y — соответственно а-, b- и у-катенины; В — винкулин; а — а-актинин; АФ — пучки актиновых микрофиламентов.
Кадхеринсодержащий адгезионный контакт является кальцийзависимым. Его регуляция осуществляется разными путями. Некоторые факторы роста, вызывая активацию определенных транскрипционных факторов, приводят к изменению уровня кадхе-рина в клетке. Факторы роста, кроме того, фосфори-лируя связанные с кадхерином белки (например, р-катенин или плакоглобин), регулируют стабильность контакта.
Наконец, G-белки семейства Rho, в частности Rac или Rho, контролирующие перестройки актинового цитоскелета, также влияют на формирование и стабильность адгезионных контактов.
Как и фокальные контакты клеток с внеклеточным матриксом, межклеточные адгезионные контакты осуществляют не только механическую связь, но также являются трансдукторами внутриклеточных сигналов, регулирующих пролиферацию клеток. Примером такой регуляции является феномен контактного торможения размножения: нормальные клетки прекращают размножаться при достижении такой плотности клеточного монослоя, когда на поверхности матрикса не остается свободных участков. Если часть монослоя клеток механически удалить, то клетки, оказавшиеся на краю «раны», начинают пролиферировать и мигрировать в свободное пространство «раны».
Таким образом, представляется весьма вероятным, что установившиеся межклеточные контакты тормозят размножение клеток.
Каковы механизмы адгезивных взаимодействий клеток друг с другом?
В сигнальной цепи, связывающей межклеточные адгезионные контакты с размножением клеток, особая роль принадлежит b-катенину. Последний находится в клетке в двух состояниях: в связанном с кадхерином (в межклеточных контактах) и в свободном (в цитоплазме). Свободный р-катенин является ключевым звеном в сигнальной цепи, функционируя как фактор транскрипции: в цитоплазме он связывается с другим транскрипционным фактором, образующийся комплекс переходит в клеточное ядро, где активирует многие гены, в том числе побуждающие клетку к делению.
Сигнальная активность свободного b-катенина и его связывание с кадхерином в адгезионном контакте конкурируют друг с другом: уменьшение связывания с кадхерином увеличивает количество цитоплазматического р-катенина, стимулируя пролиферацию клеток. Динамическое равновесие между связанным и свободным состояниями b-катенина поддерживается системой, которая регулирует в цитоплазме уровень деградации свободного b-катенина. В этой системе важную роль играют продукты генов — опухолевых супрессоров АРС и аксин.
Связываясь с цитоплазматическим b-катенином, они делают его мишенью для фосфорилирования специальной киназой, что приводит к его деградации. Возможно, что формирование межклеточных адгезивных контактов сдвигает динамическое равновесие между связанным и свободным b-катенином в сторону связанного и тем тормозит пролиферацию.
В основе контактного торможения размножения лежит также активация ряда генов — опухолевых супрессоров. Образование межклеточных контактов инициирует внутриклеточный сигнал (компоненты этой сигнальной цепи пока не изучены в деталях), вызывающий повышение экспрессии опухолевых супрессоров INK4a и KIP1, продукты которых блокируют вхождение клеток в S-фазу цикла. Продукт другого опухолевого супрессора — ТР53 накапливается в эпителиальных клетках при образовании ими сплошного пласта, приводя к остановке клеточного цикла; при разобщении клеток белок ТР53 дестабилизируется и размножение клеток возобновляется.
Важную роль играет Е-кадхерин, тоже являющийся продуктом опухолевого супрессора. Он инициирует передачу сигналов, активирующих KIP1 и ТР53, поэтому его экспрессия ведет к остановке размножения эпителиальных клеток, установивших между собой стабильные контакты.
- Вернуться в раздел "онкология"
Оглавление темы "Адгезия клеток":- Микротрубочки - структура, функции
- Стадии распластывания нормальных клеток - механизмы
- Особенности распластывания трансформированных опухолевых клеток - механизмы
- Функции актинового цитоскелета и фокальных контактов нормальных клеток
- Актиновый цитоскелет и фокальные контакты трансформированных опухолевых клеток
- Локомоция клеток - стадии, механизмы
- Особенности локомоции трансформированных опухолевых клеток
- Топографические реакции клеток - виды
- Особенности топографических реакций трансформированных опухолевых клеток
- Адгезионные взаимодействия клеток друг с другом - механизмы