Онтогенез Т-лимфоцитов - образование
Начальные (антигеннезависимые) стадии дифференцировки Т-лимфоцитов после рождения человека протекают в костном мозге, а затем в тимусе.
Первые клетки-предшественники, экспрессирующие CD7, обнаруживаются в печени плода человека на 8-й неделе внутриутробного развития. Часть этих CD7+ относится к протимоцитам, они характеризуются цитоплазматической экспрессией CD3s. На 9-й неделе протимоциты колонизируют тимус, в структуре которого присутствуют субкапсулярная, кортикальная и медулллярная области.
Появление молекулы CD7 на стволовых (CD34+) гемопоэтических клетках костного мозга является первым указанием на возможность Т-клеточной направленности их дифференцировки. Коэкспрессия CD7 и CD34 в Т-клеточном ряду дифференцировки наблюдается в течение очень короткого периода. На клетках может появляться мембранная молекула CD33. Т-клеточные предшественники, экспрессирующие только CD7 или CD7 в сочетании с CD34 (другие Т-линейные маркеры отсутствуют), обозначают как про-Т1. В Т-клеточных предшественниках костного мозга не происходит перестройки генов а- и b-цепей TCR. Фенотипической особенностью этих клеток является экспрессия CD44 при отсутствии CD25. Практически одновременно с CD7 появляется цитоплазматическая экспрессия CD3 (c-CD3). Цитоплазматическая экспрессия CD3s выявляется практически во всех случаях Т-клеточных лейкозов.
Следует помнить, что все СD3-положительные предшественники Т-клеток экспрессируют CD7 (про-Т2), в то время как из числа CD7-положительных предшественников Т-клеток только 85 % имеют C-CD3. На этом же или очень близком этапе дифференцировки на Т-клетках появляется экспрессия мембранных CD5. При ОЛЛ и лимфомах из Т-клеточных предшественников нередким является иммунологический фенотип CD7+ CD5+/- (все клетки имеют CD7, но лишь часть слабо экспрессирует CD5). На ранних этапах Т-лимфоцитопоэза (костно-мозговых, тимических) может отмечаться экспрессия молекулы CD10. Незрелые протимоциты экспрессируют CD45, но не имеют молекул HLA-II, TdT и CD3.
Рецепторный комплекс Т-лимфоцитов состоит из собственного Т-клеточного рецептора (TCR) и молекул, передающих сигнал с TCR внутрь клетки.
Т-клеточные рецепторы могут существовать в двух гетеродимерных формах — ab (90—95 % периферических Т-клеток) и уq (примерно 5—10 % периферических Т-клеток). Каждая цепь состоит из 2 доменов — константного и вариабельного, причем вариабельность выражена значительно больше, чем в V-областях иммуноглобулина. Только при объединении обеих цепей рецептор приобретает функциональную специфичность.
Передача сигнала, ведущего к активации Т-клеток после связывания TCR с презентируемым антигеном, осуществляется комплексом CD3. В составе CD3 имеется три полипептидные цепи: у, 5, s. Кроме того, с CD3 ассоциирован димер молекул e,, не относящихся к CD3. Цитоплазматические домены CD3 и e, способны взаимодействовать с белковыми тирозинкиназами после стимуляции TCR и, таким образом, передавать сигнал внутрь клетки.
Т-клеточный рецептор, CD3 и сигнальные двухцепочечные молекулы (ee) образуют Т-клеточный рецепторный комплекс.
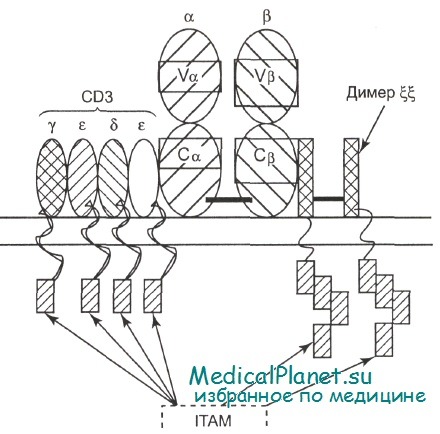
а- и b-цепи рецептора; CD3 — трансдуцерная молекула, состоящая из четырех полипептидных цепей: у, е, 8, е; димер ee; IТАМ — иммунотирози новая активационная последовательность трансдуцерных молекул.
Гены, кодирующие полипептидные цепи Т-клеточных рецепторов а/b и у/q, подобно иммуноглобулиновым генам, собираются из отдельных сегментов — V-D-J-C или V-J-C. Процесс перестройки TCRy6 начинается наиболее рано в онтогенезе Т-лимфоцитов, по-видимому, на этапе костномозговых протимоцитов. у-Ген формируется из Vy (существует 15 сегментов Vy , 10 из них подвергаются перестройке, но только 8 являются функциональными: V2_5, V8-11), Jy (3 локуса для Jyl и 2 локуса для Jy2), а также Cyl и Су2. TCRq-генные последовательности располагаются между Vа- и Ja-генными кластерами. Охарактеризованы шесть различных функциональных Vq-генов, четыре D-сегмента, три J-сегмента и один С-ген. Количество разных комбинаторных вариантов TCR уq составляет 1013.
Незрелые предшественники Т-лимфоцитов из костного мозга с током крови поступают в тимус. Становление репертуара Т-лимфоцитов происходит в тимусе. В механизмах интратимического созревания и селекции Т-клеток активную роль играют процессы генной рекомбинации и клеточного апоптоза. В тимусе формируется набор специфичностей Т-лимфоцитов, участвующих в эффекторных реакциях в периферических лимфоидных тканях.
Тимус является лимфоидно-эпителиальным органом. Основной структурный элемент тимуса — тимическая долька. Процессы внутритимической дифференцировки и селекции происходят в разных отделах дольки. В тимусе представлен широкий спектр лимфоидных и эпителиальных клеток: субкапсулярные тимические бласты, кортикальные тимоциты, медуллярные тимоциты, тельца Гассаля, дендритные клетки и т. д.
Иммунофенотипические и генетические изменения в процессе дифференцировки TCRa/b Т-лимфоцитов
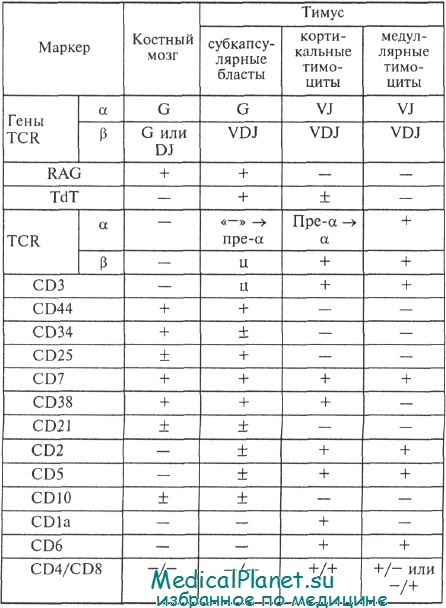
Тимоцитарная пролиферация предшествует экспрессии TCRab. При поступлении в тимус протимоциты становятся коммитированными по Т-клеточной линии и начинают экспрессировать CD3 в цитоплазме (cCD3) и TdT. Эти незрелые тимоциты являются крупными бластами и клетками среднего размера с высокими уровнями экспрессии TdT и самой большой пролиферативной активностью среди всех тимоцитов. Они являются негативными в отношении CD4, CD8 («двойные негативы»), а также CD1a. Перестройка b-генов TCR завершается на данной стадии, они еще не функционируют, белок экспрессирован только в цитоплазме. Более 90% cCD3+ незрелых тимоцитов экспрессирует рецептор ИЛ-2 (CD25). На следующем этапе дифференцировки обычные (common), или кортикальные, тимоциты являются cCD3+ (экспрессия мембранных CD3 крайне слабая), TdT. Они имеют средний уровень пролиферативной активности и отличаются по появлению мембранной экспрессии TCRp в комплексе с пре-TCRa.
В кортикальных тимоцитах происходит перестройка ос-генов, и полноценный TCRap только начинает экспрессироваться.
Мембранные пре-TCRa/TCRp и TCRa/b-гетеродимеры ассоциированы с молекулами CD3y, CD3e, CD3e и CD3q, которые имеют в цитоплазме иммунотирозиновые активационные последовательности (ITAM). Сигнал с npe-TCR через ITAM свидетельствует об эффективной перестройке и экспрессии р-генов, что необходимо для перехода тимоцитов от стадии CD4~CD8~ в стадию кортикальных тимоцитов. Эти клетки имеют на мембране CD4 и CD8 («двойные позитивы»), CD1a+, CD2+. На поздних стадиях тимического развития зрелые (медуллярные) тимоциты экспрессируют либо CD4, либо CD8, мембранные CD3+ (mCD3+), CD2+, TCRap+; TdT и CD1 утрачиваются (TdT0, CD1a0). Для перехода кортикальных тимоцитов в медуллярные необходима передача сигнала в цитоплазму клеток через полноценные TCRap, указывающая на эффективную перестройку и экспрессию а-генов.
Многие данные относительно генов TCR получены экспериментально и, возможно, у человека еще будут уточнены. Последовательность перестройки генов TCR, по всей видимости, такова: 5, у, b, а. Продуктивная перестройка TCRp на одной хромосоме ведет к блокаде рекомбинации TCRp-локуса на другой хромосоме (аллельное исключение). TCRa-гены не характеризуются аллельным исключением, и в Т-лимфоцитах человека может быть два продуктивных TCRa-гена. а-wепь собирается из 3 сегментов (V-J-C), а и-цепь — из 4 (V-D-J-C). Существуют 44 — 100 генов Va, 50—61 ген Ja и ген Са. Для р-цепей это разнообразие также очень выражено: 50—65 Vp-сегментов, 2 D-сегмента, 12—13 J-сегментов. При объединении а- и b-полипептидов может образоваться до 10" разных комбинаций.
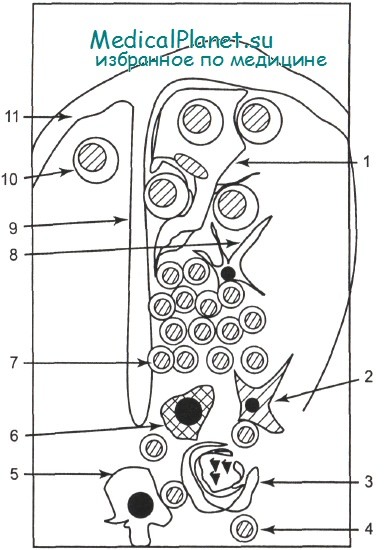
1 — клетка-нянька; 2 — интердигитирующая клетка; 3 —тельце Гассаля; 4 — медуллярный тимоцит; 5 — медуллярные эпителиальные клетки; 6 — макрофаг; 7 — агрегаты кортикальных тимоцитов; 8 — кортикальная дендритная эпителиальная клетка; 9 — септа; 10 — лимфобласт; 11 — капсула.
Разделение на линии аb и уb может происходить во время или после перестройки TCRb на уровне регуляторных участков генов TCR. Во время перестройки TCRa-гена TCR5-ген делегируется, так как он расположен между Va и Ja генными сегментами.
Из огромного числа образованных в тимусе Т-лимфоцитов, несущих разные специфичности ab-рецепторов, незначительная часть отбирается для последующего поступления в периферические лимфоидные органы, а остальные Т-лимфоциты гибнут. Позитивной тимической селекции подвергаются те CD4+CD8+TCRab+ тимоциты, которые потенциально полезны для организма. Они толерантны к аутологичным антигенам и способны распознавать чужеродные антигены в комплексе с собственными молекулами HLA на эпителиальных клетках тимуса, причем взамодействие с собственными HLA осуществляется с достаточно низким аффинитетом.
Клоны тимоцитов, которые не взаимодействуют или сильно взаимодействуют с собственными молекулами HLA, элиминируются в тимусе (отрицательная селекция). Тимоциты, распознавшие молекулу HLA-I, теряют CD4 и сохраняют CD8, а клетки, распознавшие молекулу HLA II, класса, сохраняют CD4 и теряют CD8. Тимоциты, распознающие собственные антигены, делетируются (удаляются). Это процесс отрицательной селекции, он осуществляется при взаимодействии тимоцитов с дендритными или интердигитирующими клетками. Важную роль в процессах внутритимического созревания тимоцитов играют адгезионные взаимодействия посредством молекул VLA-4, VLA-3, VLA-6, а также взаимодействия рецепторов CD2 и CD11а с соответствующими лигандами CD58 и CD54.
Для ранних этапов дифференцировки тимоцитов необходимы цитокиновые сигналы. При нарушениях в системе передачи сигнала с цитокиновых рецепторов наблюдаются тяжелые иммунодефициты. Наиболее типичным примером является тяжелая Х-сцепленная комбинированная иммунологическая недостаточность, при которой отмечается практически полное отсутствие зрелых Т- и ЕК-клеток. Дефект в этом случае касается у-цепей, общих для рецепторов ИЛ-2, ИЛ-4, ИЛ-7, ИЛ-9 и ИЛ-15.
Процессы селекции, действующие на TCRyq-клетки, отличаются от селекции, известной для TCRab-клеток, так как для уq-клеток нет HLA-ограничения.
Тимоциты, экспрессирующие TCR и прошедшие этапы интратимической селекции, дифференцируются в функционально зрелые CD4+ (Т-хелперы) или CD8+ (Т-цитотоксические) популяции. В периферические лимфоидные органы из тимуса поступают наивные Т-лимфоциты (CD4+CD8" или CD8+CD4"). Характерной иммунофенотипической особенностью этих клеток является мембранная экспрессия молекулы CD45RA, в то время как молекулы CD45RO, характерные для зрелых Т-клеток, отсутствуют.
Таким образом, тимус является центральным органом иммунной системы, ответственным за образование пула наивных Т-лимфоцитов. Позитивно отобранные клетки покидают тимус и циркулируют в крови в качестве длительно живущих покоящихся лимфоцитов. Их реактивация может произойти, как правило, только под влиянием чужеродных пептидов, представленных собственными молекулами HLA.
- Читать "Созревание (онтогенез) NK-клеток"
Оглавление темы "Клетки крови":- Онтогенез Т-лимфоцитов - образование
- Созревание (онтогенез) NK-клеток
- Созревание (онтогенез) лимфоидных дендритных клеток Лангерганса
- Рециркуляция лимфоцитов в организме
- Лимфатические узлы - формирование Т-лимфоцитов
- Функции Т-лимфоцитов лимфатических узлов
- В-лимфоциты лимфатических узлов - созревание, функции
- Внутрифолликулярная дифференцировка В-лимфоцитов
- Селезенка - строение, формирование Т- и В-лимфоцитов
- Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге