Селезенка - строение, формирование Т- и В-лимфоцитов
Селезенка является одним из важнейших периферических органов иммунной системы. Неиммунная функция селезенки заключается в разрушении и утилизации погибающих или поврежденных форменных элементов крови, а также циркулирующих микроорганизмов. Селезенка — это мощный фильтр крови и барьер для чужеродных веществ на пути из магистрального сосуда (аорты) в систему воротной вены. Как периферический орган иммунной системы селезенка участвует в иммунном ответе на растворимые антигены, циркулирующие в крови, селекции, активации и развитии толерантности В-лимфоцитов.
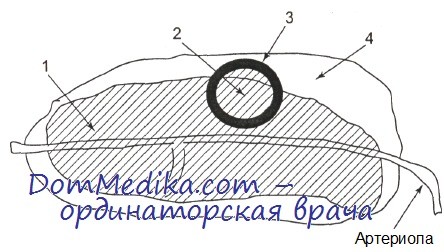
Лимфатическая ткань селезенки имеет достаточно сложное анатомическое строение. Она представлена белой пульпой и лимфоидными элементами красной пульпы. Белая пульпа сосредоточена вдоль артериол селезенки и состоит из периартериолярных лимфоидных муфт, ассоциированных с ними лимфоидных фолликулов, эллипсоидных макрофагально-лимфо-идных муфт, располагающихся на концах ветвления артериальных сосудов, и венозных синусов, в которые впадают артериолы и истинные капилляры селезенки.
Схематично строение белой пульпы селезенки представлено на рисунке. В пределах периартериолярных лимфоидных муфт (ПЛМ) выделяют наружную и внутреннюю части. ПЛМ можно считать аналогом паракортикальной (тимусзависимой) зоны лимфатических узлов. Фолликулы представляют собой В-клеточные образования. Кроме того, в селезенке имеется особая популяция В-клеток, которые отграничивают белую пульпу (периартериолярные лимфоидные муфты, включающие первичные и вторичные фолликулы) от красной пульпы. Эта область получила название краевой, или маргинальной, зоны.
Т-клетки селезенки
В селезенке присутствуют только периферические (наивные и зрелые) Т-лимфоциты, прошедшие селекцию в тимусе. Под влиянием антигенного стимула эти клетки активируются подобно тому, как это происходит в лимфатических узлах.
В белой пульпе селезенки (в периартериолярных лимфоидных муфтах) CD4+ Т-клетки преобладают, CD8+ Т-клеток меньше, а в красной пульпе наблюдается обратное соотношение между этими популяциями.
Т-клетки, имеющие рецептор TCRyq, предпочтительно оседают в синусоидах селезенки, в то время как лимфоциты, несущие TCRab, заселяют главным образом периартериолярные лимфоидные муфты.
В-клетки селезенки
В селезенке происходят процессы активации В-клеток в ходе первичных и вторичных иммунных ответов.
При формировании В-клеточного репертуара зрелые девственные В-лимфоциты поступают из костного мозга с током крови в Т-клеточные области периартериолярных лимфоидных муфт и далее в первичные фолликулы. В-клетки, специфичные в отношении аутологичных антигенов, не поступают в фолликулы, они задерживаются в наружной зоне ПЛМ и гибнут. Прошедшие фолликул В-клетки поступают в пул наивных В-лимфоцитов. Часть из них в дальнейшем активируется в селезенке, часть — в лимфатических узлах или других периферических лимфоидных органах.
Движение наивных В-клеток, поступающих из крови в селезенку, приостанавливается в наружной зоне ПЛМ. Это универсальное явление происходит после взаимодействия мембранных иммуноглобулинов В-лимфоцитов с антигеном. Биологический смысл процесса состоит в необходимости накопления активированных, пролиферирующих В-лимфоцитов в наружной зоне ПЛМ (маргинальная зона) в течение первых дней иммунного ответа на растворимые антигены белковой природы. Это позволяет многократно повысить вероятность встречи активированных наивных В-лимфоцитов с наивными Т-хелперными (CD4+) клетками той же антигенной специфичности, количество которых чрезвычайно мало.
Без Т-клеточной помощи, необходимой для реализации гуморального иммунного ответа на тимусзависимые антигены, активированные В-клетки погибают. При наличии Т-клеточной помощи наивные В-клетки поступают в фолликулы, где подвергаются соматическим гипермутациям и дифференцировке с изотипическим переключением в зародышевых центрах в ходе первичных иммунных ответов. При вторичных иммунных ответах В-клеток памяти на тимуезависимые антигены наблюдаются выраженная В-клеточная пролиферация и дифференцировка в плазматические клетки в пределах наружной зоны ПЛМ; фолликулярная В-клеточная пролиферация является несколько более слабой, чем при первичных ответах.
Антигенспецифические Т-клетки уничтожают в наружных зонах ПЛМ определенную часть В-лимфоцитов. Лиганд Fas-антигена, экспрессированный на этих Т-хелперах, включает программу гибели (апоптоз) в В-клетках с несвязанными антигенными рецепторами, а также в В-клетках, не имеющих нормального пути внутриклеточного проведения антигенного сигнала.

В тимуснезависимых иммунных ответах В-клетки способны дифференцироваться в плазмоциты без Т-клеточной помощи. При ответе на TI-антигены (ЛПС) происходят выраженная антигенспецифическая В-клеточная пролиферация и плазмоклеточная дифференцировка в наружной зоне ПЛМ и в красной пульпе; фолликулярная В-клеточная пролиферация умеренная. Считают, что именно поликлональные активаторы типа ТЫ, а также аутологичные антигены ведут к индукции CD5 на В-лимфоцитах.
CD5+ В-клетки обычно не проходят через светлый центр и не подвергаются изотипическому переключению. Интересно существование некоторого сходства CD5-лимфоцитов с толерантными аутореактивными В-клетками, которые в наружных зонах ПЛМ подвергаются абортивной активации, препятствующей их дальнейшей миграции в фолликул.
В TI-2-ответах (фиколл) большинство пролиферирующих В-клеток в наружной зоне ПЛМ дифференцируется в плазматические клетки. В селезенке существует особая область, окружающая периартериолярные лимфоидные муфты, так называемая краевая (маргинальная) зона. Клетки этой зоны являются нормальным эквивалентом клеток, из которых возникают лимфомы маргинальной зоны, или лимфоцитомы селезенки. В маргинальной зоне селезенки реализуются В-клеточные иммунные ответы на тимуснезависимые антигены, циркулирующие в периферической крови. В-клетки маргинальной зоны имеют специфические морфологические и иммунологические черты. На мембране В-лимфоцитов маргинальной зоны селезенки экспрессированы IgM, но отсутствуют IgD. Эти клетки не являются рециркулирующими, специализированы к иммунному ответу на тимуснезависимые углеводные антигены. Вместе с тем эти лимфоциты происходят из пула рециркулирующих В-клеток, возвратившихся в маргинальную зону селезенки (хоминг).
Клетки маргинальной зоны расположены в сети первичных кровяных синусоидов селезенки, что позволяет им взаимодействовать с антигенами, переносимыми кровью. Популяция клеток маргинальной зоны не столь гомогенна, как принято считать. Сюда входят В-клетки памяти, генерированные как в ходе Т-зависимых, так и Т-независимых (I тип) антительных ответов, а также девственные В-клетки, которые еще не подверглись пролиферации, зависящей от присутствия антигена. Клетки маргинальной зоны способны in vivo отвечать на все типы антигенов: Т-зависимые, Т-независимые I и II типов. Характеристики этих клеток указывают на то, что они покоятся в фазе G1, а не G0.
Ответ in vivo на TI-2-антигены (но не на Til-антигены) сильно повреждается при спленэктомии, что подтверждает отсутствие способности к рециркуляции клеток маргинальной зоны селезенки.
Их иммунофенотип сходен с иммунофенотипом большинства рециркулирующих В-клеток, но есть и некоторые отличия. Обобщенно иммунофенотипические черты В-клеток маргинальной зоны селезенки можно представить следующим образом: sIgM+ slgD- CD21+ CD35+ CD23- CD76+/- CD75-/+ CD78-2C9-/+ CD22+ CD37+ CD24- CD19+/- CD20+/- CD44+/-CD10.
В последние годы получены новые данные относительно роли селезенки в формировании репертуара длительно живущих наивных В-лимфоцитов. Из числа ежедневно продуцируемых костным мозгом ~ 1010 девственных В-лимфоцитов примерно 10% переходят в длительно живущий пул. Девственные В-лим-фоциты IgM+ IgD+ CD23- CD5- поступают первоначально в селезенку. Через концевые ветви центральных артериол и синусы маргинальной зоны они попадают в наружную зону ПЛМ. Далее небольшая часть В-клеток поступает в фолликулы. Именно эти клетки переходят в долгоживущий рециркулирующий пул. Положительная селекция В-лимфоцитов осуществляется на основании экспрессии IgVH-генов в ответ на низкие дозы экзогенных антигенов или на элементы идиотипической сети (например, сывороточные иммуноглобулины). В процессе перехода девственных клеток в наивные длительно живущие рециркулирующие лимфоциты не происходит соматических гипермутаций и переключения классов иммуноглобулинов. Пока не совсем понятно, в чем состоят иммунофенотипические различия между девственными и наивными В-лимфоцитами. Девственные В-клетки, не подвергнувшиеся положительной селекции и не поступившие в фолликулы, погибают в наружной зоне ПЛМ.
- Читать "Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге"
Оглавление темы "Клетки крови":- Онтогенез Т-лимфоцитов - образование
- Созревание (онтогенез) NK-клеток
- Созревание (онтогенез) лимфоидных дендритных клеток Лангерганса
- Рециркуляция лимфоцитов в организме
- Лимфатические узлы - формирование Т-лимфоцитов
- Функции Т-лимфоцитов лимфатических узлов
- В-лимфоциты лимфатических узлов - созревание, функции
- Внутрифолликулярная дифференцировка В-лимфоцитов
- Селезенка - строение, формирование Т- и В-лимфоцитов
- Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге