Теломераза - синтез, функции
Известно, что РНК-компонент теломеразы — hTR (human telomerase RNA) синтезируется в большинстве клеток человека независимо от теломеразной активности. Для гена белковой субъединицы теломеразы с 1998 г. принято обозначение hTERT (human telomerase reverse transcriptase). Ген hTERT состоит из 16 экзонов и 15 интронов. Общая длина hTERT составляет 37 т.п.н.. В нормальных клетках hTERT, наиболее вероятно, кодируется единичной копией гена. hTERT картирован на самом конце короткого плеча хромосомы 5 - 5р15.33. В иммортальных клеточных линиях часто наблюдают амплификацию hTERT. Видимо, амплификация hTERT— это один из способов повышения теломеразной активности в раковых клетках.
Несколько групп исследователей показали, что мРНК транскрипт hTERТ содержит по крайней мере 6 сайтов альтернативного сплайсинга. Существует тканеспецифическое распределение различных сплайсоформ hTERT в ходе онтогенеза. Это указывает на то, что сплайсинг hTERT идет неслучайным образом и, возможно, тканеспецифическирегулирует уровень теломеразной активности. Недавно проведенные исследования показали, что изменения в сплайсинге могут происходить не только в ходе развития, но и в процессе онкогенеза. Различные формы сплайсинга hTERT присутствуют в первичных образцах раковых клеток.
При анализе 5'-регуляторного района hTERT выявлено общее высокое содержание GC. Это свойство, а также наличие сайтов связывания для чувствительных к метилированию факторов транскрипции в пределах hTERT-промотора указывают на возможную роль метилирования в регуляции экспрессии hTERT. Ясно, что метилирование не ответственно за репрессию hTERT 'в большинстве соматических клеток. Многие линии раковых клеток, экспрессирующих hTERT, имеют гиперметилированные промоторные районы hTERT, а гиперметилирование самого гена hTERT во многих случаях коррелирует с теломеразной активностью.
Промоторный район hTERT богат сайтами связывания факторов транскрипции. Присутствие сайтов для многочисленных активаторов и репрессоров предполагает очень сложную систему регуляции экспрессии гена. Первым идентифицированным репрессором регуляторного района hTERT был продукт гена, супрессирующего опухоли Вильмса (WT1), который может подавлять экспрессию hTERT в некоторых клетках.
Среди факторов транскрипции, известных как активаторы hTERT, наиболее хорошо изучены онкобелок с-Мус, эстроген и прогестерон. Антагонистический контроль генной экспрессии с-Мус и Madl, вероятно, является главным регуляторным механизмом экспрессии hTERT.
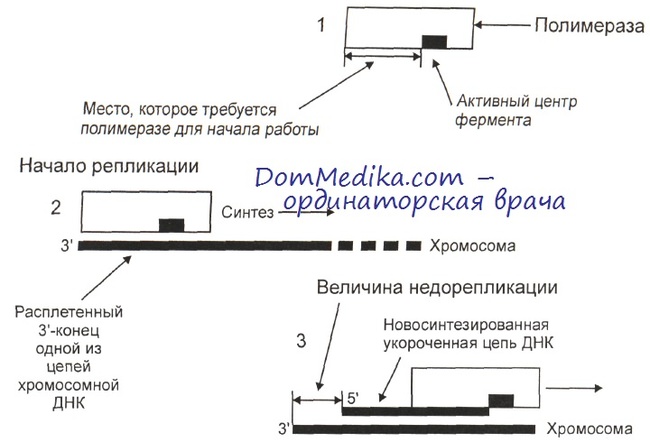
Недорепликация возникает только при репликации 3'-конца. Синтез нуклеиновых кислот может идти только в направлении 5' -> 3', поэтому репликация 5'-конца начинается изнутри хромосомы и происходит полностью. Значит, при репликации одной теломеры образуются одна точная копия и одна укороченная, которые в дальнейшем, после митоза, попадают в разные дочерние клетки.
Сборка, локализация теломер и доступ к ним. Синтез белка hTERT является основным звеном в формировании активной теломеразы у человека. Сборка активной теломеразы — энергозависимый процесс, который может включать повышение уровня транскрипции hTR, его стабилизацию и связывание с hTERT. Важным моментом, определяющим эффективную работу теломеразы, является правильная локализация этого комплекса на тело-мере. Некоторые элементы, которые участвуют в сборке активной теломеразы, известны, это белки р23 и hsp90, обеспечивающие правильное сворачивание белкового компонента. Однако остается непонятным, происходит ли полная сборка теломеразы в цитоплазме клетки, требуя, таким образом, последующего транспорта крупного комплекса в ядро, или сборка активного фермента происходит в пределах ядра.
В последнее время появилась информация о том, что теломераза, вероятно, функционирует в клетке как димер с двумя hTERT и hTR.
Посттрансляционные модификациии белка hTERT. Известно, что фосфатазы и протеинкиназы могут влиять на теломеразную активность. При этом фосфорилированию может подвергаться непосредственно каталитическая субъединица теломеразы. Установлено, что hTERT подвергается фосфорилированию киназой с-АЫ в ответ на ионизирующее облучение, причем фосфорилирование определенных специфических остатков снижает теломеразную активность.
Участие фосфорилирования hTERT в регуляции теломеразной активности было показано на гладкомышечных клетках сосудов. Гипоксия приводит к увеличению фосфорилирования hTERT, повышению теломеразной активности, но не вызывает изменений количества hTERT. Повышение теломеразной активности, по-видимому, обусловливает увеличение пролиферативного потенциала этих клеток при гипоксии.
В другом исследовании было установлено, что стимуляция лимфоцитов ведет только к незначительному увеличению количества hTERT, присутствующего в нестимулированных клетках, но вызывает значительное повышение уровня фосфорилирования hTERT, транслокацию его в ядро и возрастание теломеразной активности.
Регуляция теломеразной активности в клеточном цикле, при дифференцировке и в онтогенезе. Считается, что соматические клетки лишены теломеразной активности. Их теломеры укорачиваются как в процессе онтогенеза, так и при культивировании клеток in vitro.
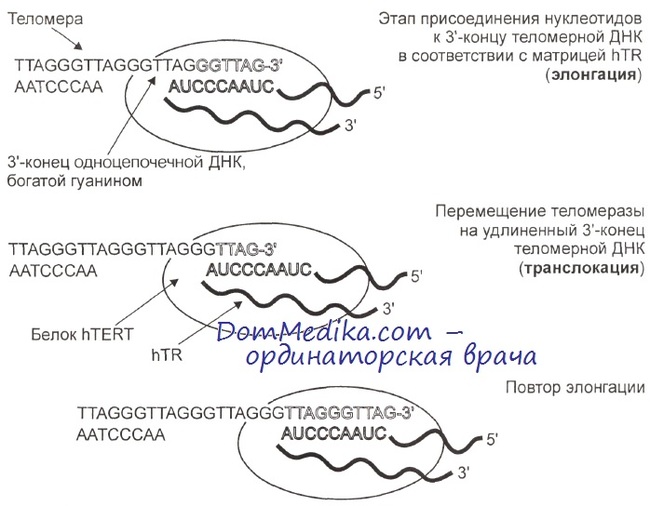
Противоречивые данные о наличии или отсутствии теломеразной активности в нормальных тканях человека и об изменениях теломеразной активности в клеточном цикле, которые появлялись в середине 90-х годов XX в., объясняются развитием и совершенствованием методов измерения теломеразной активности.
Экспериментальные данные свидельствуют, что в клетках, индуцированных к терминальной дифференцировке, может существовать несколько механизмов подавления теломеразной активности: подавление транскрипции гена hTERT, альтернативный сплайсинг, уменьшение стабильности или нарушение сборки фермента; посттрансляционные модификации, которые изменяют общую активность или сродство фермента к теломерам.
Стволовые клетки из костного мозга взрослого человека имеют более короткие теломеры, чем кроветворные клетки печени плода. Тем не менее теломеразная активность обнаруживается в нормальном костном мозге и лейкоцитах различных типов. В лейкоцитах уровни теломеразной активности варьируют, составляя 1—40 % от активности контрольных раковых клеток HeLa. При антигенной стимуляции в незрелых лимфоцитах появляется теломеразная активность, которая не обнаруживается в зрелых клетках. Установлено, что невысокие уровни теломеразной активности имеются в базальном слое эпидермиса в дифференцирующихся, временно делящихся кератиноцитах и отсутствуют в ороговевшем слое. В волосяных фолликулах, специализированных придатках кожи, наблюдается аналогичное распределение теломеразной активности в зависимости от пролиферативного состояния фолликула.
В кишечнике предполагаемые стволовые клетки расположены в нижней трети кишечных крипт. Потомство этих клеток теряет пролиферативные способности и приобретает дифференцированные свойства по мере миграции вдоль оси крипт. Теломеразная активность локализована в нижней трети кишечных крипт и не определяется в их верхней половине.
Пролиферирующие сперматоциты сохраняют значительную теломеразную активность на протяжении всей жизни. Хотя теломеразная активность присутствует в эмбриональных и взрослых тестикулах, она не детектируется в зрелых сперматозоидах, что находится в соответствии с их длительно покоящимся состоянием.
- Читать "Теломера - молекулярная структура"
Оглавление темы "Онкогенез":- Метилирование генов-супрессоров как причина гемобластозов - опухолей крови
- Теломераза - синтез, функции
- Теломера - молекулярная структура
- Что такое теломерный эффект положения?
- Альтернативные способы удлинения теломер у человека - иммортализация
- Значение теломеразы в диагностике опухолей
- Методы лечения рака влиянием на теломеры и теломеразу
- Теломеризация клеток - не ведет к злокачественной трансформации
- Адгезия клеток - последствия нарушения адгезивных взаимодействий
- Актиновые микрофиламенты - структура, функции