Тирозинкиназы и G-белки в системе передачи сигналов
Различные представители этой группы киназ вовлечены в процессы, регулирующие рост и дифференцировку клеток.
Основные категории тирозинкиназ разделены на две группы: рецепторные и нерецепторные.
Нерецепторные тирозинкиназы являются промежуточными проводниками различных внутриклеточных сигналов. Многие из этих киназ ассоциированы с трансмембранными рецепторами, например рецепторами гормонов и факторов роста, цитокинами и др.
Как и в случае стимуляции сигнала PDGF, ассоциация лиганда с рецепторными тирозинкиназами индуцирует их димеризацию либо олигомеризацию. Эти конформационные изменения приводят к индукции тирозинкиназной активности цитоплазматической области данного белка. Конформационные изменения белка играют решающую роль и в инициации нерецепторных тирозинкиназ.
Активация как рецепторных, так и нерецепторных тирозинкиназ индуцирует каскад процессов фосфорилирования (аутофосфорилирование, фосфорилирование клеточных субстратов) и вовлечение SH2-доменсодержащих белков. В результате этого происходит последовательная активизация ряда белков и вторичных проводников сигнала, которые выполняют специфичные регуляторные функции. Эти сигнальные импульсы достигают клеточного ядра и могут индуцировать экспрессию определенных генов.
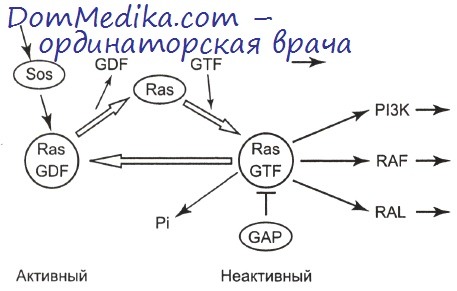
G-белки в системе передачи внутриклеточных сигналов
Белки этой группы кодируются большим семейством генов и характеризуются сходством строения и функций. Внутри семейства G-белков различают 5 больших групп: Ras, Rho/Rac, Rab, Ran и Rad/Gem, которые играют ключевую роль в процессах пролиферации, дифференцировки, изменения структуры цитоскелета, транспорта белков через ядерную мембрану и др..
Общим свойством G-белков является их способность связываться с ГДФ и ГТФ, что определило их название (GTF-proteins). В норме эти белки постоянно переходят от неактивной, ГДФ-связанной формы, к активной форме, когда они вступают в комплекс с ГТФ. Процесс переключения G-белков с ГДФ на ГТФ стимулируется «обменным» фактором (белок Sos). В этом случае активированный G-белок способен взаимодействовать со следующими молекулами (эффекторами) в определенной цепи передачи сигнала.
Антагонистами «обменного» фактора по своим функциям являются GAP-белки, которые, катализируя гидролиз ГТФ (превращение в ГДФ) в комплексе с G-белком, переводят его в пассивную форму. Следует отметить, что активность самих GAP-белков контролируется фосфорилированием под действием тирозинкиназ различных типов. Фактически малые G-белки, например ras, являются распределителями сигналов в клетке. Эти белки интегрируют получаемые от «вышестоящих» молекул (компонентов различных сигнальных путей) импульсы и активизируют последующие эффекторные белки, каждый из которых представляет собой начальное звено следующего этапа определенной цепи передачи сигнала.
Нахождение G-белков на перекрестке различных путей передачи сигналов в клетке определяет их важную роль в реализации многих происходящих в ней процессов. Нарушение функций этих белков может привести к фатальным последствиям для организма, в частности к трансформации клетки. Этим объясняется высокая частота нарушений, например протоонкогенов RAS в различных опухолях.
Превращение протоонкогенов RAS в онкогены происходит в результате действия различных молекулярных механизмов, в первую очередь вследствие мутаций в первичной структуре.
- Читать "Ядерные протоонкогены и факторы транскрипции в системе передачи сигналов"
Оглавление темы "Интерфероны. Онкогены":- Антипролиферативное действие интерферонов - механизмы
- Влияние интерферона на дифференцировку клеток
- Побочные эффекты лечения интерферонами - осложнения
- Лечение заболеваний интерферонами - применение ИФН
- Общая характеристика онкогенов
- Онкогены в системе передачи сигналов стимулированных PDGF
- Тирозинкиназы и G-белки в системе передачи сигналов
- Ядерные протоонкогены и факторы транскрипции в системе передачи сигналов
- Супрессоры опухолевого роста - антионкогены. Классификация
- Характеристика гена PML (promyelocytic leukemia gene) - функции