Единицы транскрипции иммуноглобулинов. Трансляция легких и тяжелых цепей иммуноглобулинов
Надо специально отметить, что в единицу транскрипции входят сегменты обоих константных генов, кодирующих С-концевые последовательности как секрстируемых форм IgM и IgD, так и мембранассоциированных форм этих белков. При считывании всей этой единицы транскрипции вначале образуется гигантская пре-мРНК. Сплайсинг ее различных копий может идти двумя путями.
В одном случае сегмент, кодирующий Vн-район, соединяется с сегментом, кодирующим константный район IgM, встраивающегося в мембрану лимфоцита. В другом случае с тем же сегментом мРНК для Vн-района соединяется сегмент, кодирующий участок мРНК для мембранассоциированной формы IgM. Аналогично происходит сплайсинг для секретируемых иммуноглобулинов различных классов, но одинаковой специфичности. В ходе сплайсинга вырезаются копии многочисленных интронирующих последовательностей, а также последовательностей, соответствующих исключаемому константному гену.
«Зрелые» мРНК для секретируемого и мембранассоциируемого IgM образованы соответственно 2400 и 2700 основаниями. Отличия упомянутых мРНК заключаются в 3'-концевых участках, кодирующих С-концы соответствующих u-цепей. Так, у секретируемой формы С-конец u-цепи образован 20 аминокислотными остатками, расположенными сразу вслед за Си4-доменом. Также расположен С-концевой участок мембранассоциированного IgM, но структура его совершенно иная.
На поверхности клеток иммунологической памяти, относящихся к лимфоцитам В-ряда, способны экспрессироваться одновременно идентичные по специфичности рецепторы двух классов, но в различных сочетаниях — IgM, IgG, IgM, IgE и др. Закономерен вопрос, как возникает единица транскрипции, включающая общий Vh-ген, Сu- и Са-гены, если два последних отделены друг от друга всеми остальными константными генами?
Представляется, что возникновению такой единицы предшествуют две серии реарранжировок ДНК: рекомбинация Vh-, D- и J-сегментов и деления части интрона между V- и Сu-генами, а после этого (или одновременно) S—S-рекомбинация между Сq- и СЕ-генами с последующей делецией этого участка генома. Вероятно, два процесса разобщены во времени, так что второй протекает в процессе антигензависимой дифференцировки В-лимфоцитов в клетки иммунологической памяти.
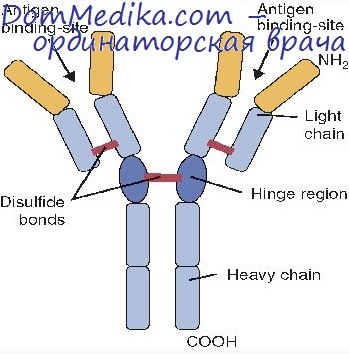
Трансляция легких и тяжелых цепей иммуноглобулинов
В зрелой плазматической клетке, синтезирующей иммуноглобулины определенного класса (изотипа), существует два вида полирибосом: одни — с мРНК для легких цепей, а другие — для тяжелых. Их скорость седиментации составляет 200S и 300S соответственно и позволяет разделить их с помощью градиентного центрифугирования. Полирибосомы для легких цепей состоят из 4—5, а для тяжелых цепей — из 11—18 рибосом. Время трансляции составляет для легкой цепи 1 мин, а для тяжелой — 2 мин.
В мРНК для легкой цепи находятся два участка, которые не транслируются. Это участок из 150 нуклеотидов на 5'-конце, который предшествует сегменту, кодирующему VL-район, и участок из 200 нуклеотидов в 3'-концсвой области между сегментом, кодирующим Cl-район, и «липким» 3'-концевым полиаденилатом.
Появляющаяся после трансляции тюлипептидная цепь отличается от легкой цепи, входящей в состав молекулы иммуноглобулина, большей молекулярной массой за счет дополнительной аминокислотной последовательности из 15—20 остатков в аминоконцевой части молекулы. Этот гидрофобный отрезок цепи необходим, по-видимому, для ассоциации полирибосомного комплекса, на котором синтезируется цепь, с мембраной эндоплазматического ретикулума, на наружной поверхности которого фиксирована полирибосома.
Указанный отрезок предшественника легкой цепи отщепляется в результате его процессинга в просвете цистерн эндоплазматического ретикулума, куда поступает новообразованный предшественник. Помимо протеолитического отщепления N-конце-вого экстрапептида именно в цистернах окончательно формируется третичная структура легкой цепи и замыкаются внутрицепьевые дисульфидные связи.
- Читать "Синтез иммуноглобулинов. Секреция новообразованных иммуноглобулинов"
Оглавление темы "Синтез и функции иммуноглобулинов":1. Рекомбинантные гены иммуноглобулинов. Гены тяжелых цепей иммуноглобулинов
2. Процессинг пре-мРНК. Процессинг иммуноглобулинов
3. Единицы транскрипции иммуноглобулинов. Трансляция легких и тяжелых цепей иммуноглобулинов
4. Синтез иммуноглобулинов. Секреция новообразованных иммуноглобулинов
5. Функции иммуноглобулинов. Антигеннезависимые эффекторные функции иммуноглобулинов
6. Реабсорбция в извитых канальцах почек иммуноглобулинов. Причина быстрого выведения Fab-фрагментов иммуноглобулинов
7. Фиксация иммуноглобулинов на тучных клетках и базофилах. Связывание иммуноглобулинов с тучными клетками
8. Цитотропные свойства IgE. Гомоцитотропины
9. Дифференцировка пре-В-лимфоцитов. Гомоцитотропины со свойствами IgG
10. Фиксация иммуноглобулинов на макрофагах (моноцитах). Связывание иммуноглобулинов с лимфоцитами