Транскрипция аденовирусов. Механизмы транскрипции аденовирусов
Аденовирус — удобная модель для изучения транскрипции и регуляции работы вирусных генов. Химические и физические свойства вируса и его ДНК хорошо известны, причем вирусную ДНК легко получить в миллиграммовых количествах. Вирус синтезирует в больших количествах мРНК, а структурные полипептиды вириона подробно охарактеризованы. Более того, с помощью аденовирусов транскрипцию можно изучать как при продуктивной инфекции, так и в трансформированных вирусом клетках. Ядра клеток, зараженных аденовирусами, синтезируют in vitro высокомолекулярную вирус-специфическую РНК.
Поскольку синтез подавляется а-аманитином, кажется вероятным, что аденовирусная ДНК в основном транскрибируется ферментом, который идентичен или очень сходен с клеточной РНК-полимеразой II (Прайс и Пенмен, 1972).
Вирусную мРНК можно разделить на два особых класса: раннюю РНК, образующуюся до начала синтеза ДНК, и позднюю РНК, появляющуюся в значительных количествах только после начала синтеза вирусной ДНК- Как следует из результатов конкурентной гибридизации, ранние и поздние мРНК качественно различны; суммарная ядерная мРНК аденовируса типа 2 имеет 58—59% ГЦ-содержания, в то время как ранняя цитоплазматическая вирусная мРНК — только 53%, что свидетельствует об избирательной транскрипции участков вирусной ДНК с относительно низким ГЦ-содержанием на ранних стадиях инфекции (Парсонс и Грин, 1971; Парсонс и др., 1971; Грин и др., 1970).
Ранние вирус-специфические РНК, которые появляются в течение 2 ч после начала инфекции, гомологичны приблизительно 8—20% вирусного генома и, будучи выделены из цитоплазмы, разделяются при электрофорезе в полиакриламидном геле на два основных компонента (23S и 16S). Если клетки заражены в присутствии циклогексимида, происходит пятикратное возрастание скорости синтеза ранних мРНК и, кроме того, становится легко определяемой вирусная мРНК еще одного вида (27S), обычно присутствующая в очень малых количествах.
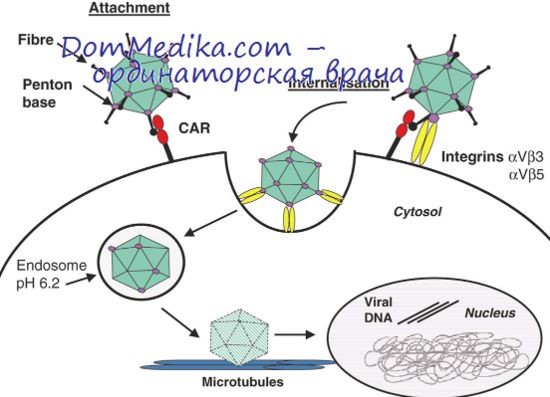
Все ранние вирус-специфические последовательности РНК определяются и на поздних сроках инфекции (через 18 ч после заражения). В трансформированных аденовирусом клетках транскрибируемые последовательности гомологичны приблизительно 50% ранних последовательностей, транскрибируемых при продуктивной инфекции (Грин и др., 1970). С этим значением хорошо согласуются результаты Рашки и Штроля (1972), которые обнаружили, что при абортивной инфекции клеток ВНК, вызванной аденовирусом типа 12, транскрибируется 60% ранних вирусных последовательностей.
Ни в трансформированных клетках, ни при абортивной инфекции в клетках ВНК поздние вирусные последовательности не были обнаружены.
Вирус-специфические РНК, которые синтезируются в клетках, трансформированных аденовирусом типа 2, гетерогенны по размеру (4—45S). 4S-PHK гибридизуется с вирусной ДНК, но, по-видимому, это не тРНК (Клайн и др., 1972). По некоторым данным, она синтезируется с помощью нечувствительной к аманитину полимеразы I, а не является продуктом распада вирусной мРНК (Прайс и Пенмен, 1972). Молекула 45S-PHK представляет собой транскрипт 30—50% вирусного генома и, по-видимому, является полицистроннои вирусной РНК. Она может содержать транскрипты клеточной ДНК, например в случае SV40.
Аденовирусную ДНК удалось разделить на фракции, содержащие тяжелую (Т) и легкую (Л) цепи (Грин и др., 1970). При продуктивной инфекции транскрибируется практически весь вирусный геном: 80% транскриптов образуется на Л-цепях и 20% — на Т-цепях; 40% ранних последовательностей мРНК являются транскриптами Т-цепей и 60%—транскриптами Л-цепей.
К поздним вирусным РНК, появляющимся через 18 ч после заражения и зависящим от синтеза ДНК, относятся обнаруживаемые в цитоплазме РНК 6—8 видов с коэффициентами седиментации в диапазоне 10—29S; размер менее крупных молекул позволяет предположить, что они служат моноцистронными матрицами. При помощи импульсной метки (15 мин) ядерной вирус-специфической РНК 3Н-уридином обнаружено четыре вида РНК (36S, 37S, 40S и 43S).
Поскольку эти меченые РНК в результате инкубации с немеченым уридином (чейза) переходили в цитоплазму, где обнаруживались меченые 10—29S-PHK, и так как последовательности вирусной РНК, присутствующие в 36—435-молекулах, методом конкурентной гибридизации были обнаружены также в цитоплазматических РНК, можно сделать вывод, что по крайней мере часть молекул цитоплазматических вирус-специфических РНК образуется при расщеплении высокомолекулярных ядерных предшественников. Поздние мРНК, определяющие синтез структурных белков вириона, относительно стабильны, в присутствии актиномицина D их количество падает, а период полужизни становится равным приблизительно 6 ч (Уайт и др., 1969).
В настоящее время быстро накапливаются данные, подтверждающие представление о том, что вслед за образованием аденовирусной РНК в виде больших молекул и их расщеплением происходит присоединение к ним полиадениловой кислоты и транспорт в цитоплазму (Филипсон и др., 1971; Раскас, 1971; Раскас и Окубо, 1971).
- Читать далее "Трансляция аденовирусов. Этапы трансляции аденовирусов"
Оглавление темы "Паповавирусы. Аденовирусы":1. Цикл размножения паповавирусов. Транскрипция паповавирусов
2. Трансляция паповавирусов. Репликация ДНК паповавирусов
3. Интеграция клеточного и вирусного геномов. Интегративность паповавирусов
4. Сборка и выход паповавирусов из клетки. Метаболизм зараженных паповавирусами клеток
5. Цикл размножения аденовирусов. Начальные стадии аденовирусной инфекции
6. Транскрипция аденовирусов. Механизмы транскрипции аденовирусов
7. Трансляция аденовирусов. Этапы трансляции аденовирусов
8. Репликация аденовирусной ДНК. Сборка и выход аденовируса из клетки
9. Изменение метаболизма инфицированного аденовирусом клетки. Цикл размножения герпесвирусов
10. Начальные стадии герпесвирусной инфекции. Транскрипция герпесвирусов