Метаболизм клеток пораженных пикорнавирусами. Синтез белков пораженными пикорнавирусами клетками
Полиовирус и другие пикорнавирусы вызывают значительное подавление синтезов РНК, белка и ДНК клетки-хозяина. Это подавление, которое начинает проявляться на весьма ранних стадиях вирусного цикла размножения, не следует путать с более поздним падением клеточных и вирусных макромолекулярных синтезов, что в конечном счете сопровождает гибель клетки при любой цитоцидной инфекции. Индуцированное вирусом подавление клеточных синтезов («shutdown», «shutoff», «cutoff») было обнаружено много лет назад (Мартин и др., 1961; Франклин и Балтимор, 1962; Циммерман и др., 1963), однако тонкие механизмы, обусловливающие этот процесс, все еще неясны (см. обзоры: Дарнел и др., 1967; Мартин и Керр, 1968).
Прямое сравнение результатов, полученных на разных системах, затруднительно, так как скорость и степень подавления очень сильно зависят как от множественности инфекции и «ростовой» (или «поддерживающей») среды, так и от штамма изучаемых вируса и клетки. Уже ранние данные о том, что клеточный белковый синтез падает по крайней мере так же быстро, как и синтез клеточной РНК, хотя мРНК в клетках животных весьма стабильна и ее период полужизни составляет несколько часов, указывали на раздельное влияние вирусной инфекции на процессы транскрипции и трансляции. Предполагается, что падение синтеза ДНК клетки-хозяина — последствие подавления синтеза белков в ней.
Большая часть имеющихся данных позволяет предположить, что подавление всех синтезов макромолекул клетки связано с вновь синтезированным белком (белками), закодированным в вирусном геноме (Франклин и Балтимор, 1962; Пенмен и Саммерс, 1965), однако вызывает недоумение то обстоятельство, что интерферон не влияет на этот процесс (Мартин и Керр, 1968).
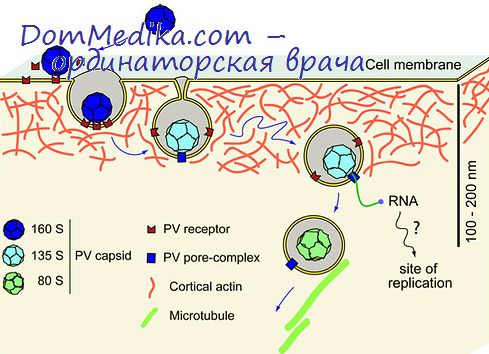
Подавление синтеза клеточных белков. На первом часу после заражения клеток пикорнавирусами полисомы начинают распадаться, и в течение 2 ч большая часть рибосом высвобождается.
Параллельно с возрастанием вирус-специфического синтеза полисомы снова появляются, но теперь они крупные и однородные по размеру (20—40 рибосом, в среднем 35), так как вРНК с мол весом 2,6 • 106 занимает место клеточных мРНК, которые гетерогенны, но в основном имеют более низкий молекулярный вес (Пенмен и др., 1963; Дейлс и др., 1965а).
К тому времени, когда в клетке появляются значительные количества новых вирусных белков, скорость клеточного белкового синтеза падает до 25—50% контроля и продолжает снижаться до конца цикла (Франклин и Балтимор, 1962). Опыты с циклогексимидом показывают, что подавление вызывается новосинтезированным белком, кодируемым вирусом (Балтимор и Франклин, 1963; Пенмен и Саммерс, 1965), который предотвращает связывание клеточной мРНК с рибосомами (Лейбовиц и Пенмен, 1971).
При заражении клеток вирусом, инактивированным ультрафолетом или гидроксиламином, а также в присутствии гуанидина репликация вирусной РНК предотвращается; в этих случаях угнетение клеточного синтеза минимально при низкой множественности инфекции, но оно становится более сильным при высокой множественности, возможно в связи с тем, что на родительских вРНК-матрицах в некотором количестве, хотя и с очень низкой скоростью, синтезируется репрессорный белок (Пенмен и Саммерс, 1965). Купер и др. ( 1973) предполагают, что репрессор — это «эквестрон», т. е. вирусный капсидный белок-предшественник NCVP1 на той стадии, когда он расщепился па VP0, 1 и 3, но перед его конечным расщеплением на VP1, 2, 3 и 4.
Эренфелд и Хаит (1971) показали, что двухцепочечная РНК (РФ) полиовируса подавляет синтез белка in vitro в лизатах ретикулоцитов кролика. Следует отметить, что это явление неспецифично, так как синтетические полинуклеотиды и двухцепочечная РНК фага Q|3 или реовируса также обладают сходным иигибирующим действием.
- Читать далее "Подавление синтеза клеточных РНК пикорнавирусами. Подавление синтеза ДНК пикорнавирусами"
Оглавление темы "Характеристика тогавирусов и ортомиксовирусов":1. Метаболизм клеток пораженных пикорнавирусами. Синтез белков пораженными пикорнавирусами клетками
2. Подавление синтеза клеточных РНК пикорнавирусами. Подавление синтеза ДНК пикорнавирусами
3. Дефектные частицы пикорнавирусов. Цикл размножения вируса ящура и тогавирусов
4. Трансляция тогавирусов. Этапы трансляции тогавирусов
5. Репликация РНК тогавирусов. Транскрипция РНК тогавирусов
6. Сборка и выход тогавирусов из клетки. Метаболизм пораженной тогавирусами клетки и флавивирусы
7. Размножение вируса краснухи в клеточных культурах. Цикл размножения ортомиксовирусов
8. Начало ортомиксовирусной инфекции. Транскрипция ортомиксовирусов
9. Трансляция ортомиксовирусов. Репликация РНК ортомиксовирусов
10. Сборка ортомиксовирусов. Выход ортомиксовирусов из клетки