Ранняя стадия парамиксовирусной инфекции. Транскрипция парамиксовирусов
Адсорбцию парамиксовирусов на чувствительных клетках детально не исследовали, однако присутствие гемагглютинина и нейраминидазы во внешней вирусной липопротеидной оболочке дает основания предположить, что процесс в основном не отличается от аналогичного процесса у ортомиксовирусов. Вопрос о механизмах проникновения и раздевания парамиксовирусов, так же как и в случае других вирусов с липопротеидной оболочкой, вызывает споры. Компанс и др. (1966) получили электронные микрофотографии, демонстрирующие примеры цитоза SV5, тогда как Дьюранд и др. (1970) утверждают, что высвобождение нуклеопротеида ВНБ происходит после слияния вирусной оболочки с изолированными мембранами куриных клеток.
Хомма (1971) описал интересный пример зависимой от хозяина модификации парамиксовируса, который имеет отношение к пониманию процесса проникновения. Вирус Сендай проходит в L-клетках только один цикл, его потомство неинфекционно для L-клеток (но инфекционно для куриных эмбрионов), а гемолитическая активность сильно снижена. При обработке такого выращенного в L-клетках вируса трипсином в низкой концентрации его инфекционность для L-клеток повышается в 1000 раз и существенно восстанавливается гемолитическая активность. Подобным же образом большой урожай высокоинфекционного вируса со значительным содержанием гемолизина можно получить, обработав трипсином сами L-клетки в то время, когда происходит почкование нового вируса.
Показано, что трипсин удаляет с поверхности выращенного на L-клетках вируса Сендай белок, который в противном случае мешает присоединению вирусной оболочки к плазматической мембране. Эти данные находятся в согласии с гипотезой о том, что парамиксовирусы проникают в клетку в результате слияния вирусной и клеточной оболочек с помощью гемолизина, действие которого связано либо с лизолецитином вирусной оболочки (Барбанти-Бродано и др., 1971), либо с ее пепломерами (Хосака и Симизу, 1972).
Определенный интерес представляют также данные о том, что размножение вируса Сендай блокируется строфантином G, который, как известно, подавляет фермент плазматической мембраны (Na, К)-АТФазу; такое подавление приводит к неспособности клеток поддерживать высокую внутриклеточную концентрацию ионов К+ (Нагаи и др., 1972).
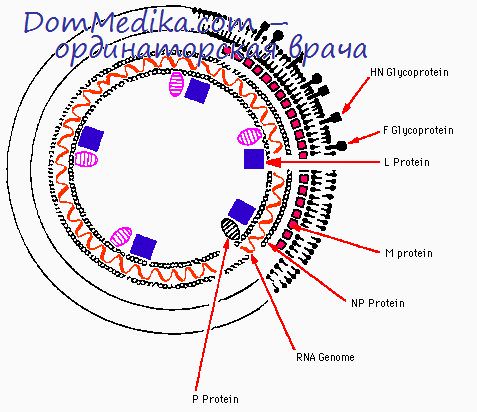
Транскрипция парамиксовирусов
РНК, синтезирующаяся в клетках, зараженных ВНБ, в основном представляет собой одноцепочечную кРНК, которая на основании размера м,ожет быть отнесена к двум гетерогенным классам, 18S и 35S; РНК обоих классов связывается с полисомами (Кингсбери, 1966; Брэт и Робинсон, 1967). Подобные данные получены и для вируса Сендай (Блэр и Робинсон, 1967). Поэтому кажется вероятным, что мРНК парамиксовирусов состоит из молекул кРНК с меньшим размером, чем вРНК-матрица (50—57S), на которой они синтезируются. Вслед за транскрипцией к 18S-KPHK ковалентно присоединяется полиадениловая кислота, имеющая коэффициент седиментации приблизительно 4S (Приджен и Кингсбери, 1972).
У вируса Сендай транскрипция родительской вРНК-матрицы с образованием кРНК происходит и в присутствии циклогексимида (Робинсон, 1971), вероятно потому, что в клетку вместе с нуклеокапсидом вносится вирусная транскриптаза (Хуанг и др., 1971; Робинсон, 1971а; Стоун и др., 1971). На поздних стадиях цикла циклогексимид также не влияет на синтез кРНК, которая в это время все еще является основным видом образующейся РНК, однако этот ингибитор быстро подавляет синтез вРНК (57S) (Робинсон, 1971b).
Портнер и Кингсбери (1972) выделили из клеток, зараженных вирусом Сендай, структуру, которую они назвали транскриптивным предшественником (ТП). Он представляет собой быстрометящуюся гетерогенную (28—60S) РНК с двухцепочечными участками и одноцепочечными хвостами, что подтверждается его частичной устойчивостью к РНКазе и способностью осаждаться в 1 М NaCl.
Эта структура при плавлении дает смесь 50S- и 18S-PHK фракция была идентифицирована как вРНК, а 18S-фракция (путем гибридизации с выделенной из вирионов РНК)—как кРНК; эту интерпретацию нужно принимать с некоторой осторожностью, так как было показано, что небольшая часть 50—57S-PHK, выделенная из очищенных вирионов ВНБ или вируса Сендай, комплементарна основному виду РНК—вРНК (Робинсон, 1970).
Вероятно, из некоторых ТП должны образовываться молекулы кРНК полной длины (50—57S), которые в свою очередь служат матрицами для синтеза новых вРНК. Важнейший вопрос о том, каким образом в процессе транскрипции парамиксовирусной вРНК обеспечивается синтез как длинных, так и коротких кРНК-транскриптов, остается неразрешенным. Одна из возможностей заключается в том, что все образующиеся транскрипты имеют полную длину, но впоследствии некоторые из них расщепляются, вероятно в несколько стадий, образуя моноцистронные матрицы.
В противном случае необходимо предположить, что транскриптаза, по-видимому отличная от известной вирионной транскриптазы, обладает способностью считывать вРНК не останавливаясь в точках пунктуации. Выяснение этой интересной проблемы имеет значение отнюдь не для одних парамиксовирусов.
Транскриптаза, связанная с нуклеокапсидом вириона (Хуанг и др., 1971; Робинсон, 1971а; Стоун и др., 1971), возможно, является РНК-зависимой РНК-полимеразой, которая количественно обнаруживается в микросомной фракции цитоплазмы на поздних стадиях цикла репликации (Мейхи и др., 1970). Активность как одного, так и другого фермента зависит от Mg2+ и четырех рибонуклеозидтрифосфатов; их температурный оптимум 28° С, а оптимум рН 8, они не подавляются актиномицином D и катализируют in vitro образование коротких одноцепочечных кРНК
- Читать далее "Трансляция и репликация РНК парамиксовирусов. Сборка и выход парамиксовирусов из клетки"
Оглавление темы "Характеристика парамиксовирусов и рабдовирусов":1. Метаболизм клетки пораженной вирусом гриппа. Зависимость вируса гриппа от клеток хозяина
2. Неполный вирус гриппа. Парамиксовирусы
3. Ранняя стадия парамиксовирусной инфекции. Транскрипция парамиксовирусов
4. Трансляция и репликация РНК парамиксовирусов. Сборка и выход парамиксовирусов из клетки
5. Метаболизм пораженных парамиксовирусами клеток. Слияние клеток пораженных парамиксовирусами
6. Размножение коронавирусов человека. Размножение аренавирусов и супергруппы Буниамвера
7. Вирус везикулярного стоматита. Цикл размножения и транскрипция рабдовирусов
8. Трансляция рабдовирусов. Репликация РНК рабдовирусов
9. Сборка и выход рабдовирусов из клетки. Степень выраженности генов рабдовирусов
10. Метаболизм пораженных рабдовирусом клеток. T-частицы рабдовирусов