Метаболизм клеток пораженных реовирусами и орбивирусами. Генетика вирусов
Синтез клеточных макромолекул не повреждается сколько-нибудь значительно вплоть до относительно поздних стадий цикла. Синтез клеточной РНК подавляется при высокой множественности заражения. В течение первых 7 ч синтез клеточных белков не изменяется, и даже к 9 ч он падает только до 50% (Энсмингер и Тамм, 1969; Звееринк и Иоклик, 1970). Кинетика подавления синтеза клеточной ДНК подобна кинетике подавления синтеза белков (Энсмингер и Тамм, 1969).
Причина подавления синтеза клеточной ДНК, по-видимому, заключается в уменьшении числа мест инициации, а не в снижении скорости роста цепи (Хэнд и Тамм, 1972).
Вирус синего языка овец, типичный вирус этого рода (обзоры: Фервурд, 1970; Хауэл и Фервурд, 1971), размножается в клетках ВНК21 или L с латентным периодом 4 ч, достигая максимальных титров через 12 ч после начала инфекции. В отличие от реовирусов вирус синего языка очень быстро подавляет синтез белка клетки-хозяина (Хюйсманс, 1971).
Для вируса синего языка характерно такое же соответствие между мРНК и геномными фрагментами, как и для реовирусов (гл. 3). При транскрипции 10 геномных фрагментов образуется 10 видов мРНК, молярные отношения которых после синтеза in vitro или in vivo, а также после выделения из полисом говорят о некоторой специфической регуляции на уровне транскрипции, в то время как регуляция на начальных стадиях трансляции, по-видимому, отсутствует (Хюйсманс, Фервурд, 1972). Подробные сведения о молекулярной биологии размножения орбивирусов еще не получены.
Электронно-микроскопические исследования размножения нескольких орбивирусов в культуре клеток или в мозге мышей показали, что созревание осуществляется в «гранулярных» или «ретикулярных» цитоплазматических включениях, содержащих множество нитей и трубочек. Показано также, что вирус выходит из клетки не путем почкования, а в результате лизиса клетки (Мёрфи и др., 1971а).
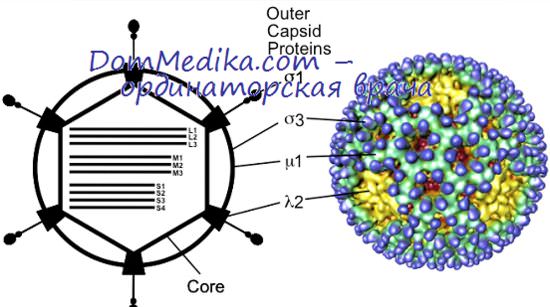
Генетика вирусов
В молекулярной генетике — наиболее плодотворной части молекулярной биологии — с самого начала и до сих пор центральное место занимает интенсивное изучение одной-единственной бактерии, Escherichia coli, и ее многочисленных вирусов-паразитов (Кэрнс и др., 1966; Хейс, 1968; Стент, 1971). Генетическое исследование вирусов животных сильно отставало— главным образом потому, что работать с диплоидными клетками позвоночных значительно труднее, чем с гаплоидными клетками Е. coli. Начало успешному генетическому изучению вирусов животных было положено, когда Дульбекко ввел метод бляшек на клетках позвоночных. Исследования в этой области все шире развертываются благодаря трем факторам: улучшенной технике работы с клеточными культурами, более разнообразным и точным методам биофизического и биохимического изучения вирусов и их компонентов и растущему использованию условно-летальных мутаций.
В статьях мы рассмотрим мутации вирусных генов и взаимодействия между вирусными геномами или их продуктами при смешанной инфекции клетки.
Взаимодействие между вирусами нельзя рассматривать как чисто экспериментальный феномен; множественная инфекция клеток позвоночных как в лаборатории, так и в природе встречается, вероятно, даже чаще, чем единичная. Многие, если не все, клетки интактного животного или клеточных культур могут быть заражены нецитоцидными вирусами (в свободной форме или в виде провируса), так что суперинфекция клеток различными вирусами (т. е. смешанная инфекция) — явление весьма обычное. В плотных органах (например, в печени), на слизистых оболочках и в клеточном монослое распространение инфекции из клетки, даже если она была заражена лишь одной вирусной частицей, обычно сопровождается множественной инфекцией соседних клеток. Результат заражения несколькими частицами зависит от того, идентичны они или принадлежат разным вирусам (или разным мутантам одного вируса).
Одновременное множественное заражение несколькими инфекционными частицами одного и того же вируса может сократить латентный период и уменьшить асинхронность процесса; это важно при биохимическом изучении репродукции вирусов. В связи с низкой эффективностью инфекции, характерной для большинства вирусов животных, большая часть вирионов при множественном заражении не в полной мере участвует в инфекции. Некоторые частицы неинфекционны, другие потенциально инфекционны, но по каким-то случайным причинам не вызывают инфекции. Однако такие частицы могут участвовать в генетическом взаимодействии и уменьшать урожай инфекционного вируса — в результате интерференции (гл. 8) или образования «неполного» вируса. Заражение одной и той же клетки разными вирусами может приводить а) к их независимой репродукции, б) к задержке размножения одного или обоих вирусов, в) к комплементации и усиленному размножению одного или обоих вирусов, г) к генетической рекомбинации, д) к фенотипическому смешиванию или е) к различным сочетаниям этих взаимодействий.
В настоящее время нет согласованной терминологии для обозначения взаимодействий между вирусами. Мы будем использовать общий термин «рекомбинация» для всех случаев обмена генетическим материалом между вирусами, происходит ли он на внутримолекулярном уровне или путем перераспределения отдельных молекул нуклеиновой кислоты; «комплементацией» мы будем называть все случаи увеличения урожая одного или обоих вирусов при смешанной инфекции клеток.
- Читать далее "Вирусные гены. Количество генов у вируса"
Оглавление темы "Лейковирусы и реовирусы":1. Циклы размножения лейковирусов. Начальная стадия лейковирусной инфекции
2. Транскрипция лейковирусов. Синтез ДНК на матрице лейковирусной РНК
3. Синтез и трансляция мРНК лейковирусов. Сборка и выход лейковирусов из клетки
4. Метаболизм клеток пораженных лейковирусами. Цикл размножения реовирусов
5. Транскрипция реовирусов. Этапы транскрипции реовирусов
6. Трансляция реовирусов. Репликация РНК реовирусов
7. Сборка реовирусов. Степень выражения генов реовирусов
8. Метаболизм клеток пораженных реовирусами и орбивирусами. Генетика вирусов
9. Вирусные гены. Количество генов у вируса
10. Исследование вирусного генома. Методы анализа вирусного генома