Лизосомы клеток. Рибосомы клеток
Лизосомы чуть мельче митохондрий и представляют собой пузырьки, окруженные единичной гладкой липопротеидной мембраной и содержащие множество гидролитических ферментов. Лизосомные мембраны образуются, вероятно, путем почкования аппарата Гольджи, в частности при контакте с ним пиноцитозных пузырьков. Лизосомы встречаются в большей части клеток позвоночных, но особенно многочисленны они в фагоцитах, где образуют гранулы, хорошо знакомые гематологам.
При фагоцитозе лизосомы высвобождают свое содержимое в фагоцитозные пузырьки, что влечет за собой быстрое переваривание захваченного клеткой материала.
От цитоплазмы клеток гидролитические ферменты надежно отделяются лизосомными мембранами. По предположению Эллисона (1967а), лизосомы могут участвовать в трех стадиях цикла размножения вирусов животных. Они могут поставлять ферменты, раздевающие вирион внутри фагоцитозного пузырька. Ферменты лизосом могут быть также вовлечены в разрушение полинуклеотидов клетки-хозяина, которое приводит к значительному увеличению фонда кислоторастворимых нуклеотидов в зараженной вирусом клетке.
И наконец, они могут играть роль в процессе клеточной дегенерации, способствуя высвобождению вирусов, созревающих во внутренних участках клетки, а не на клеточной мембране, и вызывая цитопатический эффект (ЦПЭ), используемый как показатель размножения вируса в культуре клеток.
Комплекс Гольджи представляет собой стопку уплощенных пузырьков, находящихся на одном из полюсов клетки и расположенных между ядерной оболочкой и секреторными пузырьками. Комплекс можно изолировать из клеточных гомогенатов методом дифференциального центрифугирования. Функция аппарата Гольджи точно не известна, однако вполне возможно, что именно здесь остатки сахаров присоединяются к белкам (см. обзор Ньютры и Леблонда, 1969).
Комплекс Гольджи особенно хорошо развит в секреторных клетках, в связи с чем полагают, что он принимает участие в переносе белков из эндоплазматической сети к наружной поверхности клетки.
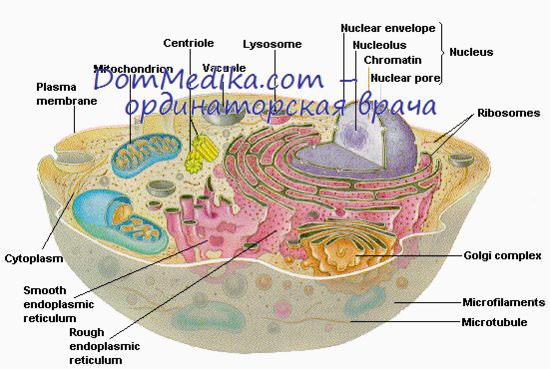
Наиболее крупные и сложные цитоплазматические органеллы клетки — митохондрии; наиболее мелкие - рибосомы, но они столь многочисленны (около 107 на одну клетку HeLa), что на их образование затрачивается до 25% общего числа макромолекул, синтезируемых в клетке. Рибосомы имеют структуру сплющенных сфероидов размером приблизительно 25X18 нм, коэффициент седиментации рибосом близок к 80 S (Спирин и Гаврилова, 1969).
При электронной микроскопии негативно контрастированных препаратов видно, что каждая рибосома состоит из двух неодинаковых субъединиц (60 S и 40 S), отделяющихся друг от друга в отсутствие ионов магния или в растворах с высокой концентрацией КСl (Жирар и др., 1965; Мартин и Вул, 1968). Каждая субъединица состоит из большого числа различных полипептидных цепей, состав и число которых до настоящего времени все еще полностью не установлены, и одной или двух молекул РНК.
60S-субъединица содержит РНК с молекулярным весом 1,75-106 дальтон, возможно, в виде двух фрагментов, связанных водородными связями. В неденатурирующих условиях эта РНК седиментирует при 28 S (Пин и др., 1968). 40S-cyбъединица содержит единственную молекулу РНК с коэффициентом седиментации 18 S и молекулярным весом 0,70-106 дальтон.
Кроме того, рибосомы содержат 5S-PHK, локализация которой неизвестна, но нуклеотидная последовательность определена (Форгет и Вайсман, 1969; Уильямсон и Браунли, 1969). Рибосомные РНК обладают рядом характерных признаков: высоким содержанием Г и Ц, большим количеством метилированных оснований, а также содержат необычные основания, например псевдоуридин.
Почти все этапы биосинтеза рибосом проходят в ядрышке, так как именно там находится так называемая рибосомная ДНК (см. обзор Бирнштиля и др., 1971). Эта ДНК включает последовательности, комплементарные 18S- и 28S-pибосомным РНК. а также спейсерные участки. ДНК содержит большое количество повторяющихся последовательностей, и методом гибридизации при насыщающих концентрациях в ДНК клеток HeLa обнаружено около 1100 копий рибосомных последовательностей (Жантёр и Аттарди, 1969).
Возможно, они образуют прилегающие друг к другу блоки, в которых чередуются последовательности, комплементарные 18S- и 28S-pPHK (см. Браун и Вебер, 1968). Рибосомная ДНК транскрибируется, по-видимому, особой ДНК-зависимой РНК-полимеразой (Блатти и др., 1970), в результате чего образуются молекулы РНК с коэффициентом седиментации 45 S и молекулярным весом 4,1*106 дальтон.
Эта РНК служит предшественником рибосомных РНК; она последовательно и специфично расщепляется на несколько фрагментов, два из которых в конечном счете представляют собой рибосомные 18S- и 28S-PHK. Как осуществляется это расщепление (процесоинг), неизвестно; непонятно также, каким образом сохраняется целостность рибосомных последовательностей, в то время как оставшаяся часть предшественника деградирует.
Новосинтезированная рибосомная РНК образует комплекс с рибосомными белками в ядрышке, и в цитоплазму выходят полностью сформировавшиеся 40S- и 60S-субъединицы рибосом. Несмотря на то что выделено несколько промежуточных структур, участвующих в процессе сборки рибосом, механизм морфогенеза рибосом остается неясным. Место синтеза рибосомных белков неизвестно; время от времени появлялись сообщения о том, что они синтезируются на ядерных рибосомах, однако в настоящее время представляется более вероятным, что они образуются на обычных цитоплазматических рибосомах, а затем транспортируются обратно в ядрышко.
В цитоплазме новосинтезированные 40S- и 60S-частицы включаются в фонд рибосомных субъединиц, 80S-mohocom и полисом. Полисомы состоят из групп рибосом, связанных цепью информационной РНК. Они представляют собой «рабочий станок», на котором синтезируются полипептиды. Вероятнее всего, все вирусные полипептиды синтезируются на полисомах; даже в том случае, когда вирионы собираются в ядре (как у аденовирусов), вирусные белки, по-видимому, синтезируются на полисомах в цитоплазме.
- Читать далее "Эндоплазматическая сеть клеток. Ядро и хромосомы клеток"
Оглавление темы "Клетки. Особенности деления клеток":1. Движение клеток. Контактное торможение клеток
2. Пиноцитоз клеток. Митохондрии клеток
3. Лизосомы клеток. Рибосомы клеток
4. Эндоплазматическая сеть клеток. Ядро и хромосомы клеток
5. Ядрышки клеток. Клеточный цикл
6. Синтез хромосомной ДНК. Синтез макромолекул ядра клеток
7. Синхронизация клеточного деления. Биохимия живых клеток
8. Строение хромосом. Структура хромосом
9. Транскрипция нуклеиновых кислот. Этапы транкскрипции
10. Информационная РНК. Трансляция