Эндоплазматическая сеть клеток. Ядро и хромосомы клеток
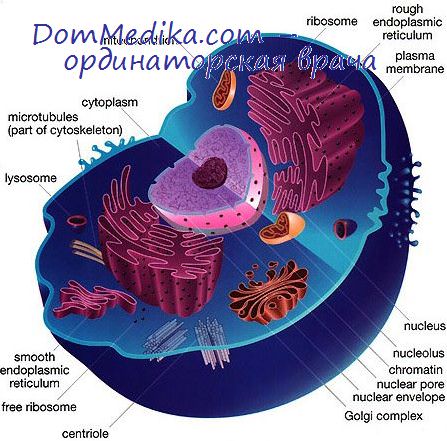
В клетках, производящих и секретирующих большие количества специфических видов белков (например, в плазматических клетках, образующих антитела, секреторных клетках поджелудочной железы и других желез), рибосомы связаны с мембранами, образуя шероховатую эндоплазматическую сеть. В большинстве клеток, используемых для культивирования вирусов, шероховатая эндоплазматическая сеть развита слабо, однако этот тип мембран никогда не отсутствует полностью даже в клетках HeLa.
Микросомная фракция клеточного гомогената состоит главным образом из рибосом и фрагментов эндоплазматической сети; полисомы в таких препаратах обычно бывают разрушены в связи с ферментативной деградацией мРНК, связывающей рибосомы.
Ядро и хромосомы клеток
В отличие от бактерий у высших организмов генетический аппарат отделен от цитоплазмы и сосредоточен в специальной клеточной органелле—ядре. Ядро окружено ядерной мембраной и содержит хромосомы и одну «ли более специализированных органелл, называемых ядрышками.
В интерфазной клетке ядро окружено оболочкой, состоящей из двух концентрических элементарных мембран, отстоящих друг от друга на 10—15 нм. Ядерная оболочка, по-видимому, представляет собой продолжение мембран эндоплазматической сети. Она пронизана расположенными через постоянные интервалы порами, где параллельные внутренняя и внешняя мембраны сливаются вокруг отверстий различного диаметра.
Поры занимают около 10% поверхности ядерной оболочки (Уотсон, 1959). С помощью методов негативного контрастирования было показано, что поры имеют восьмиугольную форму и окружены цилиндрическими структурами, называемыми колечками (Гол, 1967), которые, возможно, функционируют как диафрагма, регулирующая поступление и выход частиц из ядер.
Во время митоза ядерная оболочка исчезает, распадаясь на отдельные мелкие пузырьки; на стадии поздней телофазы она образуется вновь, как полагают, из цитоплазматической мембранной системы (Барер и др., 1959).
В диплоидных клетках млекопитающих содержится около 3,8-1012 дальтон ДНК. В культивируемых клетках в связи с их тенденцией к анеуплоидии количество ДНК обычно несколько выше. Вопросы о том, находится ли ДНК в хромосоме в виде одной или нескольких молекул, а также каков способ ее упаковки в интерфазных ядрах, остаются неразрешенными (Дьюпро, 1970).
Не подлежит сомнению, однако, что большая часть ДНК связана с основными белками, протаминами и гистонами. Протамины — это очень простая и гомогенная группа полипептидов с молекулярными весами приблизительно 4000 дальтон; гистоны крупнее и разнообразнее, их молекулярные веса лежат в пределах от 10 до 18 тыс. дальтон.
Оба вида белков в очень большом количестве содержат основные аминокислоты, аргинин и лизин и, по-видимому, присоединяются к одной из бороздок двойной спирали ДНК. Гистоны и протамины, несомненно, каким-то образом регулируют проявление генетической информации, закодированной в ДНК (Пол и Гилмор, 1968), но механизм этой регуляции остается до сих пор неизвестным.
Комплекс ДНК—белок (хроматин) в живом интерфазном ядре обычно не виден. Однако непосредственно перед, делением клетки хроматин конденсируется, образуя хромосомы, которые видны в световом микроскопе. Число хромосом и их структура — чрезвычайно характерные свойства каждого вида животных (Су и Бениршке, 1967).
Различные хромосомы, составляющие кариотип животного, в настоящее время можно безошибочно отличить друг от друга по следующим трем характеристикам. Во-первых, каждая хромосома имеет определенный размер. Во-вторых, у каждой хромосомы есть небольшая перетяжка — центромера, представляющая собой область присоединения хромосомы к митотическому веретену.
Положение центромеры в каждой хромосоме строго определенно, поэтому указанный признак можно с успехом использовать в целях классификации. В-третьих, для каждой хромосомы характерно определенное расположение полос при окрашивании по методу Гимза после щелочной или тепловой денатурации ДНК (Гол и Пардью, 1969; Пэтил и др., 1971) или при использовании хинакринового иприта (Касперсон и др., 1970).
Во время интерфазы хромосомы становятся такими длинными и тонкими, что их практически не видно, за исключением двух необычных случаев: хромосом типа ламповых щеток в некоторых ооцитах и политенных хромосом у некоторых двукрылых. Хромосомы типа ламповых щеток имеют ось, от которой отходят парные петли.
Результаты радиоавтографических исследований показали, что в петлях идет активный синтез РНК и белка (Изава и др., 1963). Активность петель различна: одни активны постоянно, другие лишь в начале и конце роста ооцита. Гол и Каллан (1962) обнаружили, что в хромосомах типа ламповых щеток из ооцитов тритона синтез РНК начинается в определенном локусе на одном конце петли и постепенно распространяется по направлению к другому (см. обзор Каллана, 1963). Прекрасные электронные микрофотографии хромосом типа ламповых щеток, на которых можно проследить этот процесс, получены Миллером и его коллегами (Миллер и др., 1970).
Пуффы политенных хромосом у различных двукрылых также вовлечены в процесс транскрипции (Беерман и Клевер, 1964; Эшбёрнер, 1970). Все сказанное наводит на мысль, что образование петель или пуффов — это проявление активности генов, расположенных в данном участке, и что дифференциальное действие генов в процессе развития регулируется, по крайней мере отчасти, механизмами, определяющими специфическую спирализацию и деспирализацию различных участков хромосомы.
- Читать далее "Ядрышки клеток. Клеточный цикл"
Оглавление темы "Клетки. Особенности деления клеток":1. Движение клеток. Контактное торможение клеток
2. Пиноцитоз клеток. Митохондрии клеток
3. Лизосомы клеток. Рибосомы клеток
4. Эндоплазматическая сеть клеток. Ядро и хромосомы клеток
5. Ядрышки клеток. Клеточный цикл
6. Синтез хромосомной ДНК. Синтез макромолекул ядра клеток
7. Синхронизация клеточного деления. Биохимия живых клеток
8. Строение хромосом. Структура хромосом
9. Транскрипция нуклеиновых кислот. Этапы транкскрипции
10. Информационная РНК. Трансляция