Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vivo
Исследования in vivo. При внутримозговом заражении мышей, как указывалось выше, развивается смертельное заболевание. К моменту гибели в организме животных обычным методом не обнаруживаются ни нейтрализующие, ни комплементсвязывающие антитела. Однако такое «иммунологическое безразличие» к вирусу ЛХМ на самом деле оказывается лишь кажущимся; ото было установлено благодаря использованию различных методов иммунодепрессии.
Так, удаление зобной железы, введение антилимфоцитарной сыворотки, лимфоцитотоксических веществ, облучение Х-лучами предотвращают летальный исход ЛХМ-инфекции (Hotchin, Lehmann-Grube). Такой эффект пе связан с изменением титра вируса, что хорошо было продемонстрировано в опытах с циклофосфамидом. Однократное введение 150 мг/кг этого препарата через 3 дня после заражения защищало от гибели 90% животных, при этом титры вируса и распределение вирусного антигена в организме обработанных и необработанных мышей были сходны (Cole, Gilden е. а.).
Следовательно, защитное действие иммунодепрессантов не связано с изменениями вирусной репродукции, а определяется иммунной реакцией инфицированного организма. Выжившие мыши получили название «лекарственно-индуцированные взрослые носители», так как в течение всей последующей жизни в их крови и других тканях определяется вирус в титре 103 ЛД50/мл, хотя и в этом случае не обнаруживаются ни комплементсвязывающие, ни нейтрализующие антитела.
Напротив, если спустя 5 дней после заражения таким «лекарственно-индуцированным взрослым носителям» ввести в брюшную полость 108 селезеночных клеток, иммунных к лимфоцитарному хориоменингиту мышей, то защитное действие циклофосфамида снимается и животные спустя 6 дней погибают при клинической картине острого лимфоцитарного хориоменингита, что в последующем подтверждают и результаты гистологического исследования.
Описанный эффект иммунологической реконструкции не удается получить при внутрибрюшинном введении тем же животным идентичного количества селезеночных клеток неиммунного животного (Cole е. а., Gilden е. а.). Не удается вызвать заболевания и введением больших объемов иммунной сыворотки. Наконец, предварительная обработка иммупных клеток селезенки анти-Т-лимфоцитарной сывороткой полностью снимает их способность к иммунологической реконструкции, но пе снимает их способность вырабатывать комплементсвязывающие антитела (Cole).
Таким образом, описанные выше опыты прямо указывают па клеточную природу иммунитета к лимфоцитарному хориоменингиту. Этот вывод подтверждается и другой серией исследований, где использовали внутрибрюшинный способ введения вируса.
В подобных случаях развивается ограниченный патологический процесс, не затрагивающий центральной нервной системы. В сыворотке крови таких животных через 8—10 дней после заражения появляются комплсментсвязывающие антитела, достигающие максимальных титров (1:50) к 14-му дню.
Заражение таких мышей в мозг дозой 1000 ЛД50 вируса ЛХМ через неделю и позднее после впутрибрюнлшного заражения показывает полную резистентность животных. При этом наблюдают ограниченную репродукцию вируса в певральных мембранах центральной нервной системы до 3-го дня с последующим снижением его титра и полным исчезновением к 10-му дню. Обнаруживаемые в это время признаки воспаления сосудистой оболочки мозга неярко выражены и никогда не достигают интенсивности, наблюдаемой у зараженных в мозг неиммунных мышей.
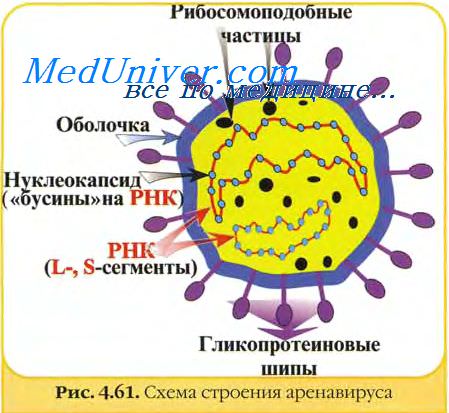
Свидетельство приобретенной невосприимчивости к внутримозговому введению вируса получено также после пересадки нормальным мышам свободных от вируса клеток иммунных животных — 109 селезеночных клеток или 108 клеток брюшного экссудата. Через несколько дней реципиенты становятся резистентными к внутримозговому заражению. У таких животных после испытания резистентности в крови накапливается высокая концентрация (1:50) комплементсвязывающих антител. Однако передача здоровым мышам таких количеств сыворотки, которые создают у них такие же или даже более высокие концентрации антител, не обеспечивает им невосприимчивости к вирусу лимфоцитарного хориоменингита.
Это, несомненно, подтверждает вывод о клеточной природе иммунитета к ЛХМ-инфекции. В данном случае такая клеточная реакция опосредует развитие смертельного заболевания центральной нервной системы, а в другом — она же защищает организм от него. Столь разная направленность действия иммунологического механизма, приводящая в разных случаях к прямо противоположным исходам, объясняется динамикой вирусной репродукции и ее взаимоотношением с сенсибилизированными лимфоидными клетками: своевременная, если так можно выразиться, мобилизационная готовность лимфоидных клеток вызывает развитие активного, но ограниченного воспалительного процесса, который успевает оборвать инфекционный процесс до того, как значительные количества клеток будут вовлечены в перестройку клинически значимой мишени для иммунологической атаки (Cole, Nathanson).
Постоянные неудачи при попытках обнаружить антитела у животных, страдающих различными формами заболевания, и у вирусоносителей привели к общему мнению о толерантности мышей к вирусу лимфоцитарного хориоменингита. На этой основе даже был предложен специальный термин — «персистентная толерантная инфекция» (Hotchin). Прошло много лет, прежде чем с помощью техники непрямого МФА в организме вирусоносителей удалось обнаружить циркулирующие вирусоспецифические антитела (Oldstone, Dixon, Benson, Hotchin).
В настоящее время считают, что развитие гломерулонефрита вызвано отложением в почечных клубочках иммунных комплексов, состоящих из вирусного антигена, комплементсвязывагощих антител и, по крайней мере, одной фракции комплемента С3. Исходя из такого представления о патогенезе медленной инфекции, вызванной вирусом лимфоцитарного хориоменингита, можно искусственно получить у носителей признаки заболевания в сроки, значительно более ранние чем через 6 мес после рождения.
Для этого проводят опыт перекрестного кормления врожденно инфицированных новорожденных мышат иммунными к вирусу лимфоцитарного хориоменингита матерями. В результате характерные для лимфоцитарного хориоменингита повреждения развиваются уже через месяц после рождения (Oldstone, Dixon).
Как уже описывалось выше, персистентная инфекция вирусом лимфоцитарного хориоменингита формируется в трех случаях:
- у мышей старшего возраста («лекарственно-индуцированные взрослые носители»);
- при заражении новорожденных;
- в результате вертикальной передачи вируса.
Адоптивная иммунизация вирусоносителей любого типа соответствующим количеством сингенных лимфоидных клеток от иммунного донора приводит к развитию патологических повреждений, характер и степень тяжести которых зависят от типа персистентной инфекции, а также от ее предварительной продолжительности (Cole е. а., Gilden е. а.). Так, адоптивная иммунизация взрослых вирусоносителей не позднее 10-го дня после инфекции приводит к развитию классической картины лимфоцитарного хориоменингита.
Если же прививку произвести позднее 30-го дня после инфицирования, то развивается менее тяжелый и часто несмертельный ЛХМ, но у таких животных обнаруживают очаговые некротические поражения мозжечка. Результат введения клеток новорожденным или врожденным носителям также зависит и от момента переноса. Если он осуществляется вскоре после инфицирования, то патологические изменения включают очаговый некроз центральной нервной системы и повреждения органов. В случае переноса клеток таким носителям, которые уже достигли взрослого состояния, у них через 5 дней развивается несмертельное генерализованное заболевание, которое продолжается около 10 дней и характеризуется очаговыми поражениями печени при полном отсутствии повреждений в центральной нервной системе (Hannover, Volkert).
К наиболее поразительному следствию адаптивной иммунизации мышей-вирусоносителей относится появление через 2—3 нед очень высоких титров антител (равных или превышающих 500), которые по своей природе могут быть и нейтрализующие, и комплементсвязывающие или только комплементсвязывающие, что зависит от штаммов вируса или мышей (Volkert, Hannover, Larsen, Volkert, Lundstedt, Gilden o. a.).
Появление антител сопровождается выраженным процессом удаления вируса из тканей и крови животных, механизм которого пока не совсем ясен. Нельзя исключить того, что этому процессу способствуют нейтрализующие антитела, тогда как комплементсвязывающие антитела не играют никакой роли, так как у адоптивно иммунизированных взрослых носителей, которые были инфицированы в новорожденном состоянии, можно наблюдать одновременное присутствие в высоких титрах и инфекционного вируса, и комплементсвязывающих антител (Lundstedt).
Кроме того, установлено, что иммунная элиминация вируса из организма мышей, поддерживающих любую форму инфекций, требует участия специфичных для вируса Т-лимфоцитов (Cole е. а., Zinkernagel, Doherty).
- Читать "Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vitro"
Оглавление темы "Медленные вирусные инфекции":- Иммунитет при врожденной краснухе
- Эпидемиология врожденной краснухи - распространенность
- Вирус лимфоцитарного хориоменингита (ЛХМ) - структура, характеристика
- Клиника лимфоцитарного хориоменингита (ЛХМ) - проявления
- Патогистология лимфоцитарного хориоменингита (ЛХМ)
- Патогенез лимфоцитарного хориоменингита (ЛХМ)
- Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vivo
- Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vitro
- Эпидемиология лимфоцитарного хориоменингита (ЛХМ) - распространенность
- Возбудитель лихорадки Ласса - структура, характеристика