Патогенез и патологическая анатомия бешенства. Чувствительность к вирусу бешенства
В патогенезе и патологической анатомии бешенства ведущим признаком является поражение всей нервной системы, в том числе нервных ганглиев некоторых железистых органов и особенно слюнных желез, через которые вирус выделяется во внешнюю среду. Патологоанатомические изменения со стороньГ головного и спинного мозга, слюнных желез, надпочечников характеризуются отечностью, полнокровием сосудов, участками кровоизлияний. При гистологическом исследовании головного мозга обнаруживают диффузный энцефалит в отличие от локальных поражений, встречающихся при других инфекциях ЦНС.
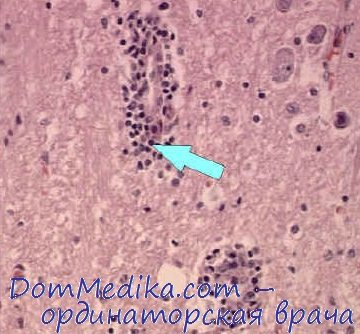
Наблюдается явное несоответствие между клинической картиной тяжелого рабического энцефалита и сравнительно скудными находками ультраструктурных изменений, отсутствием воспалительных изменений вокруг телец Бабеша — Негри, слабо выраженной воспалительной реакцией микроглии и оболочек. Сосудистая реакция выражается в кровенаполнении, расширении мелких вен и капилляров, набухании эндотелия и инфильтрации просвета элементами белой крови. Инфильтраты вокруг сосудов состоят из лимфоцитов и глии. Воспалительные гранулемы, или узелки Бабеша, располагаются поблизости измененных сосудов и представляют из себя пролиферацию глии. Нервные клетки могут изменяться по форме и в размерах (набухание, сморщивание), подвергаться зернисто-жировому перерождению и некрозу.
Некоторое диагностическое значение имеет дегенеративно-воспалительное изменение спинномозговых ганглиев и ганглиев вегетативной нервной системы. При экспериментальном бешенстве, вызванном фиксированным вирусом, отмечаются менее выраженная периваскулярная реакция и несколько более выраженные внутриклеточные изменения: оксифилыюе перерождение ядер клеток, образование вакуолей, некроз, увеличение лиаосом. В ЦНС вирус бешенства накапливается главным образом в сером веществе и очень редко в белом веществе. Специфический антиген обнаруживается чаще всего в нейронах аммонова рога, пирамидальных клетках мозжечка, нейронах других отделов головного и спинного мозга, а также в эпителиальных клетках слюнных желез, роговицы и не обнаруживается в менингеальных, эпендимальных, глиальных клетках, а также в эндотелии сосудов ЦНС (Yamamoto, Johnson). При электронно-микроскопическом исследовании Matsumoto наблюдал образование вирусного матрикса в цитоплазме астроцитов. Следовательно, селективное поражение нейронов не имеет абсолютного значения (Johnson).
Чувствительность макроорганизма к вирусу бешенства зависит от вида животного, возраста его, зоны инокуляции, от свойств штамма, а также от дозы вируса. Чрезвычайно чувствительны дикие плотоядные животные (лисица и др.), высокочувствительны сирийский хомяк, крупный рогатый скот и др., умеренно — кролик, собака и малочувствительны опоссумы. Эффективность воспроизведения болезни в эксперименте повышается при ионокуляции вируса в следующей последовательности: в просвет сосудов, брюшную полость, подкожную клетчатку, внутримышечно, внутри кожно, в ствол нерва, в мозг. В естественных условиях заражение вирусом бешенства происходит через поврежденные кожные покровы. Считается, что неповрежденные кожные покровы непроницаемы, по через неповрежденные слизистые оболочки верхних дыхательных путей и конъюктивы вирус может проникать.
В литературе описано аэрогенное заражение человека:
а) в пещере Фрио (штат Техас) с миллионами естественно инфицированных вирусом бешенства летучих мышей (Constanline) и
б) лаборанта во время центрифугирования мозгового материала, содержащего фиксированный вирус бешенства (Winkler е. а.). В условиях эксперимента при большой концентрации вируса возможно заражение лабораторных животных как оральным, так и аэрогенным путем: закапывание в нос или вдыхание аэрозолей в камере (И. А. Свет-Молдавская, М. А. Селимов и др., Atanasiu).
Установлено, что у входных ворот инфекции вирус бешенства может сохраняться до 2 нед. В отношении вируса летучих мышей допускается возможность его размножения в мышечной ткани у входных ворот инфекции (Sulkin с. а.). Большинству исследователей не удалось подтвердить размножение его па месте введения (Schindler, Dean e. a.).
Для объяснения путей проникновения вируса в ЦНС были выдвинуты различные гипотезы. Основными из них являются гипотезы о нейрогенном и гематогенном путях. Более убедительны данные экспериментов о иейрогеипом пути распространения. Margagni еще в 1789 г. писал: «...кажется, что вирус бешенства проходит в ЦНС не по венам, а по нервам» (цит. по Johnson). О нейрогенном распространении вируса бешенства свидетельствуют следующие данные: а) выделение вируса из периферических нервных стволов людей и животных, погибших от бешенства, обнаружение вируса в соответствующем периферическом нерве через несколько часов после заражения (Roiix, Di Vestea, Zagari, Nicolau-Galloway); б) при периферическом заражении животных гистологические изменения установлены в соответствующих нервных стволах и узлах (Goodpasture, 1925; Sabin е. а., Field);
в) при заражении животных в нервный ствол или интраплантарно перерезка соответствующих нервных стволов на пути продвижения вируса предупреждает развитие болезни (Nicolau, Mateiesco, Dean e. a.). Nicolau, Mateiesco подтвердили центробежное распространение этого вируса во второй фазе инфекционного процесса: у зараженного в мозг животного дистальный конец перерезанного седалищного нерва не содержит инфекционного вируса. По данным Dean с соавт., вирус бешенства в опыте на морских свинках продвигается по нерву со скоростью 3 мм в час.
В последние годы в нашей стране зарегистрированы 2 случая рождения здоровых детей от женщин, больных гидрофобией. Martell с соавт. сообщили об обнаружении вируса бешенства в мозге плода коровы, инфицированной в естественных условиях. Пока безуспешны попытки заразить экспериментальных животных вирусом бешенства через кровососущих паразитов-клещей (Bell е. а.). В опытах Schwcinburg, Windholz рабическая инфекция не переходила к партнеру-парабионту при общей циркуляции крови и отсутствии нервных связей. Zunker, повторив опыты предыдущих авторов, как будто наблюдал заболевание партнера-парабионта. Таким образом, невозможно полностью исключить возможность и гематогенного распространения вируса бешенства, особенно в отношении вируса, адаптированного к организму летучих мышей.
Вирус бешенства во время центростремительного движения проникает как в ЦНС, так и в слюнные железы. В последних он размножается не только в нервных ганглиях, но и в клетках эпителия канальцев. Еще до развития клинических признаков болезни, после достаточного накопления, начинается экскреция вируса во внешнюю среду из слюнных желез. Активное размножение вируса одновременно имеет место, по-видимому, в надпочечниках и других железистых органах (слюнные железы и др.). В большинстве внутренних органов брюшной и грудной полостей вирус не размножается, и в них могут быть обнаружены лишь следы вируса, проникшего в соответствующие нервные волокна из ЦНС во время центробежного его распространения. Все же американский вирус летучих мышей, обладающий некоторой висцеротропностью, выделяется из крови, мочи, легочной ткани, сердца и из поперечнополосатой мускулатуры больных животных.
- Читать "Иммунитет к вирусу бешенства. Естественный и поствакцинальный"
Оглавление темы "Прививки от бешенства":- Прививки от бешенства в нашей стране. Качество антирабической помощи
- Обращаемость за антирабической помощью. Источники укусов в нашей стране
- Причины прививок от бешенства в Европе и Америке. Источники бешенства
- Причины прививок от бешенства в Африке и Азии. Укусы животными
- Патогенез и патологическая анатомия бешенства. Чувствительность к вирусу бешенства
- Иммунитет к вирусу бешенства. Естественный и поствакцинальный
- Перекрестный иммунитет к вирусу бешенства. Интерферон
- Роль клеток крови в формировании иммунитета к бешенству. Нервные клетки и вирус бешенства
- Формирование антирабического иммунитета. Уровни защиты организма от бешенства
- Антирабические вакцины. История