Вирус лимфоцитарного хориоменингита (ЛХМ) - структура, характеристика
Лимфоцитарный хориоменингит (ЛХМ) — зоонозное инфекционное заболевание преимущественно мышей, характеризующееся поражением центральной нервной системы, в особенности мозговых оболочек и сосудистых сплетений, в развитии которого ведущую роль играют иммунологические механизмы. Болезнь передается людям, у которых могут развиться разнообразные ее формы. Возбудитель лимфоцитарного хориоменингита относится к группе аренавирусов.
Аренавирусы — таксономическая группа, включающая чувствительные к растворителям липидов вирусы сходной структуры, содержащие РНК и группоспецифический антиген, определяемый с помощью МФА (Rowe е. а.). Несколько необычное название группы происходит от латинского «arenaceus» (песчаный, песочный), в связи с тем что вириопы аренавирусов содержат необычные чувствительные к РНК-азе грапулы, которые хорошо видны на ультратонких срезах.
В настоящее время в группу аренавирусов, кроме вируса лимфоцитарного хориоменингита (прототипный вирус), входят также вирус лихорадки Лаоса и вирусы комплекса Такарибе, т. е. Джунин, Такарибе, Ама-пари, Латино, Мачупо, Парана, Пичинде, Тамиами.
Вирус лимфоцитарного хориоменингита был выделен в 1933 г. Armstrong и Lillil от человека, погибшего во время эпидемии энцефалита Сап-Луи. Авторы заражали испытуемым материалом обезьян с последующим пятикратным пассированием инфекционного агента из этих животных. И так как последняя обезьяна оказалась иммунной к вирусу энцефалита Сан-Луи, полагали, что вирус ЛХМ выделен от одной из обезьян, а не из тканей погибшего человека.
Traub через 2 года изолировал вирус лимфоцитарного хориоменингита от мышей, которые заболевали после внутримозгового введения им стерильного бульона, а также из крови работника вивария, который ухаживал за инфицированными мышами.
Электронно-микроскопический анализ позволил выявить значительный полиморфизм вирионов лимфоцитарного хориоменингита, характеризующихся обычно сферической, реже чашеобразной формой, размеры которых колеблются от 50 до 200 нм и более. Внутри частиц обнаружены от 1 до 8 и даже более электронно-плотных гранул, чувствительных к РНК-азе, снаружи вирионы покрыты шипиками. Формирование зрелых вирионов осуществляется отпочковыванием от плазматической мембраны клетки (Dalton е. а., Abelson е. а., Kajima).
Вирус лимфоцитарного хориоменингита чувствителен к действию детергентов, эфира (Stock е. а., Andrewes, Horstmann), мертиолята (Rogers), pH 3 (Padnos е. а.) и двухвалентных катионов (Pfau, Camyre). Напротив, 0,05—0,005 М трис-буфер оказывает стабилизирующее действие на вирус, правда, при условии содержания больших количеств белка (Pfau).
Вирус оказывается стабильным при анализе в градиенте плотности хлорида рубидия или хлорида цезия, вирусная активность обнаруживается в этих условиях в зонах с плотностью 1,15 и 1,24 г/мг (Pfau).
Изучение физико-химических свойств вируса лимфоцитарного хориоменингита, полученного через 24 ч после инфицирования клеток ВНК-21 и очищенного осаждением насыщенным раствором сульфата аммония с последующим фракционированием в градиенте сахарозы, показало, что плавучая плотность очищенного вируса соответствует 1,18 г/мл. При анализе вирусоспецифической РНК выделены 4 крупных компонента с константами седиментации 31S, 28S, 23S, 18S. Все они представляли собой одионитчатые РНК. Три других малых компонента РНК обладали констаптами седиментации 4S, 5S и 5,5S.
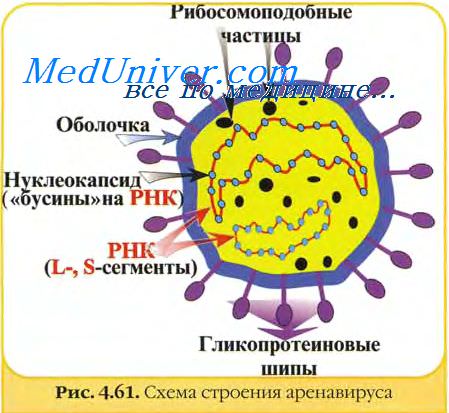
Вирусоспецифическими оказались лишь РНК 28S, 31S, остальные виды были рибосомальными клеточными РНК. Вирус лимфоцитарного хориоменингита, меченный по 3Н-валину, содержал 1 большой и 5 меньших белковых компонентов (Pedersen).
Изучение свойств другого представителя аренавирусов — вируса Пичинде дало сходные результаты. После обработки вирио-нов неионным детергентом NP-40 с вирусной РНК остаются ассоциированными 1 гликопептид и 1 полипептид. Вирусная РНК имеет однопитчатую структуру и распределяется в градиенте сульфата цезия в зоне с плотностью 1,66—1,68 г/мл, что характерно для односпиральпой РНК.
Геном вируса Пичинде сегментирован и состоит из 5 дискретных компонентов РНК — 31S, 28S, 22S, 18S и 4—6S, но анализ вирусиндуцированпой РНК из зараженных клеток, обработанных актиномицином D, показал, что фрагменты 31S и 22S РНК кодируются вирусным геномом, а фрагменты 28S, 18S и 4—6S РНК, по-видимому, клеточным (Bowls е. а., 1973).
В организме инфицированных животных или в питательной среде инфицированных клеточных культур накапливается так называемый растворимый комплементсвязывающий антиген, который может быть отделен от вирусных частиц. Сам антиген не обладает инфекционностью и может быть экстрагирован бутанолом из органов зараженного животного или из инфицировапных клеточных культур.
Такой антиген сам по себе не вызывал иммунного ответа у животных, а в смеси с адъюваптом Фрейнда индуцировал у морских свинок и кроликов синтез комплементсвязывающих антител, по пе вируснейтрализующих антител (Gschwender, Lehmann-Grube). Комплементсвязывающий антиген не защищал морских свинок от заражения вирусом ЛХМ, а аптисыворотка против комплементсвязывающего антигена не реагировала с очищеппым вирусом. Нейтрализующая активность гипериммунной антисыворотки против вируса лимфоцитарного хориоменингита не блокировалась при добавлении комплементсвязывающего антигена.
Комплементсвязывающий антиген, экстрагируемый бутанолом из инфицированных клеток и тканей, не локализуется на поверхности вирионов и неизвестно, является ли он их структурным компонентом (Gschwender, Lehmann-Grube).
Вирус лимфоцитарного хориоменингита репродуцируется в большинстве испытанных клеточных культур, в куриных эмбрионах, культурах макрофагов мыши. В культуре клеток куриных или мышиных эмбрионов, первичных культурах амниона человека, Детройт-6, KB, мышиной лимфосаркомы, L репродукция вируса сопровождается цитопатическим действием. Репродукция вируса в культурах почек и сердца обезьян 3Т3, ВНК-21, RE, МК-2, СМИ, НЕр-2, печеночных клеток Chang, а также жировой ткани не сопровождается видимыми признаками клеточной деструкции (Hotchin).
Клеточные культуры используют при изучении вируса лимфоцитарного хориоменингита не только для моделирования различных форм инфекции, но и как один из наиболее эффективных методов получения различных штаммов вируса. Так, например, из линии клеток L, инфицированных вирусом, выделили штамм, не вызывающий цитопатического эффекта. Этот штамм обладал низкой патогепностью для мышей и обеспечивал им последующую невосприимчивость к вирулентному вирусу даже при внутримозговом введении последнего (Lehmann-Crube е. а., Hotchin, Sikora).
Нередко селекция штаммов различной патогенности удавалась при использовании тоста бляшкообразования, как это имело место с культурой BHK-21/13S, зараженной субштаммами МВ8 и MB6L (суспензий мозга или печени зараженных мышей соответственно) штамма WE вируса лимфоцитарного хориоменингита. Первый из субштаммов формировал под агарозой 96,5% прозрачных бляшек, а второй — 10,1%. Субштамм МВ811 при заражении клеточного монослоя вызывал цитопатическии эффект, а при введении в головной мозг мышам — гибель животных, тогда как субштамм MB6L не вызывал клеточной дегенерации в слое, а в организме мыши приводил к развитию хронической инфекции (Hotchin е. а.).
Использование метода клеточных культур для определения инфекционного вируса не требует много времени, экономично и просто, но этот метод в отличие от метода внутримозгового заражения мышей менее чувствителен (Gschewender, Lehmann-Grube).
Вирус лимфоцитарного хориоменингита вызывает заболевание у человека, белых мышей, морских свинок, домовых мышей, крыс, резусов и макак (Hotchin). Однако он формирует бессимптомную инфекцию у цыплят, собак, кроликов, новорожденных и взрослых сирийских хомяков, у новорожденных и иногда у взрослых мышей (Hotchin).
- Читать "Клиника лимфоцитарного хориоменингита (ЛХМ) - проявления"
Оглавление темы "Медленные вирусные инфекции":- Иммунитет при врожденной краснухе
- Эпидемиология врожденной краснухи - распространенность
- Вирус лимфоцитарного хориоменингита (ЛХМ) - структура, характеристика
- Клиника лимфоцитарного хориоменингита (ЛХМ) - проявления
- Патогистология лимфоцитарного хориоменингита (ЛХМ)
- Патогенез лимфоцитарного хориоменингита (ЛХМ)
- Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vivo
- Иммунитет при лимфоцитарном хориоменингите (ЛХМ) - исследования in vitro
- Эпидемиология лимфоцитарного хориоменингита (ЛХМ) - распространенность
- Возбудитель лихорадки Ласса - структура, характеристика