1. мРНК. Промотор. РНК-полимераза. Транскрипционные факторы. Энхансер. Транскрипция мРНК. Процессинг. Интрон. Экзон.
2. Полипептидная цепь. Биосинтез полипептидной цепи. Сплайсинг. Сплайсеосомы. Транспортная РНК. тРНК.
3. Рибосомы. рРНК. Синтез полипептидной цепи на рибосоме. Рибосома и мРНК.
4. Структура гена. Тонкая структура гена. Организация гена у эукариот. Мультигенные семейства. Псевдогены.
5. Геном человека. Характеристика генома человека. Свойства генома человека. Особенности генома человека.
мРНК. Промотор. РНК-полимераза. Транскрипционные факторы. Энхансер. Транскрипция мРНК. Процессинг. Интрон. Экзон.
Для того чтобы произошло списывание последовательности нуклеотидов гена, кодирующих первичную структуру определенной полипептидной цепи белка на мРНК, к цепи ДНК на некотором расстоянии от гена, к специальной последовательности нуклеотидов, называемых промотором, присоединяется РНК-полимераза.
Этот фермент разрывает фосфорные связи между нуклеотидами ДНК в области соответствующего гена и делает обе цепи ДНК доступными для считывания. Присоединению РНК-полимеразы к промоторной области генов способствуют так называемые общие и специфические транскрипционные факторы3, которые, как и РНК-полимераза, являются продуктами других генов.
В транскрипции могут участвовать и другие белки, так что в целом процесс транскрипции является достаточно сложным и зависящим от действия многих генов. Белки, участвующие в транскрипции, в своей структуре содержат так называемые мотивы, обеспечивающие их связь с ДНК. Принципиально обе нити ДНК могут считываться, но обычно считывается только одна, и это зависит от последовательности нуклеотидов промотора.
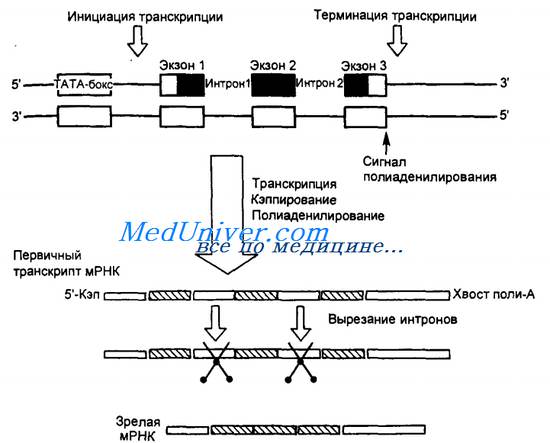
РНК-полимераза движется от 3'- к 5'-концу вдоль соответствующей нити ДНК, способствуя комплементарному присоединению нуклеотидов синтезируемой молекулы мРНК к цепи ДНК в направлении от 5'- к З'-концу (рис. 3.7).
Стартовой точкой транскрипции служит основание ДНК, соответствующее основанию РНК, которое первым включается в транскрипт. Транскрипция мРНК продолжается до тех пор, пока РНК-полимеразе II не встретится сигнал терминации транскрипции. Первичный транскрипт мРНК расщепляется специфической эндонуклеазой сразу за последовательностью нуклеотидов АААУАА и к З'-концу мРНК пришивается с помощью поли-А-полимеразы хвост из 100—200 остатков аде-нина, которые, по-видимому, защищают мРНК от деградации во время ее передвижения из ядра в цитоплазму. Кроме того, вскоре после начала синтеза мРНК ее 5'-конец «копи-руется», т.е. к нему присоединяется модифицированный нук-леотид — 5-метилцитозин. Кэппирование происходит в месте начала транскрипции. Обычно транскрипция продолжается, и мРНК удлиняется еще на несколько тысяч пар нуклеотидов, комплементарных ДНК, после чего молекула мРНК отъединяется от матрицы ДНК и РНК-полимеразы. Прежде чем такая мРНК попадет в цитоплазму, она должна созреть (см. рис. 3.7).
Созревание, или процессинг, заключается в том, что из первичного транскрипта мРНК вырезаются (сплайси-руются) специальными ферментами участки гена, транскрибированные в мРНК, которые содержат обычно (но не всегда) некодирующие последовательности1. Эти некодирующие участки имеются практически во всех генах высших организмов и называются интронами. От молекулы мРНК отрезаются также последовательности нуклеотидов за поли-А-хвос-том. Кодирующие участки гена называют экзонами. Экзоны гена, представленные в мРНК, соединяются с помощью специальных ферментов вместе, образуя функционально зрелую мРНК. Именно последовательность кодонов мРНК кодирует последовательность аминокислот в белке, который будет создаваться на ее основе. Зрелая мРНК перемещается в цитоплазму.
P.S. Промотор и прилегающие к нему участки являются последовательностями нуклеотидов ДНК, которые выполняют очень важную функцию — они должны распознаваться белками, имеющими отношение к транскрипции РНК и контролю за этим процессом; промотор генов у эукариот обычно включает ТАТА-бокс, ГЦ или ГГГЦГГГ консенсусную (т.е. эволюционно консервативную) последовательность и ЦЦААТ или ЦАТ-бокс — короткие последовательности нуклеотидов, которые распознаются РНК-полимеразой.
У эукариот три РНК-полимеразы, которые обеспечивают транскрипцию трех разных классов генов: РНК-полимераза I ответственна за транскрипцию рибосомальных генов, РНК-полимераза II — за транскрипцию генов белков и некоторых типов РНК, а РНК-полимераза III — за транскрипцию генов для остальных типов РНК.
К транскрипционным факторам относят энхансеры, которые значительно ускоряют процесс транскрипции. Для того чтобы энхансеры могли осуществить свою функцию, они должны связаться со специфическими транскрипционными факторами, которые называют активаторами. Активаторы в свою очередь связываются с коактиваторами, также специфическими, т.е. разными для разных генов. Наконец, образовавшийся комплекс связывается с неспецифическими транскрипционными факторами и с самим геном. Параллельно энхансерам, усиливающим транскрипцию, в геноме существуют последовательности ДНК, ослабляющие ее (сайленсеры). Эти последовательности также связываются с комплексом специфических и неспецифических транскрипционных факторов и геном, снижая транскрипционные возможности последних.
- Читать далее "Полипептидная цепь. Биосинтез полипептидной цепи. Сплайсинг. Сплайсеосомы. Транспортная РНК. тРНК."