Эффективность цельных ослабленных вирусов в вакцине от ВИЧ
Некоторые исследователи несколько лет назад показали, что иммунизация макак живым, ослабленным естественным путем штаммом SIV (SIVmac1A11) не защищала от инфицирования, но предотвращала развитие заболевания у взрослых и новорожденных животных при последующем введении вирулентного штамма вируса.
Кроме того, предшествовавшее инфицирование макак изолятом SHIV89,6 обеспечивало защиту при влагалищной инокуляции патогенного штамма SIVmac239. В этом случае развитие защиты, скорее всего, может быть связано с активацией перекрестных клеточных реакций, чем с выработкой нейтрализующих антител.
Кроме того, у макак, иммунизированных перорально двумя живыми ослабленными изолятами SHIV, развивалась защита от заболевания в случае, если вирус вакцины сохранялся в организме животного в активной форме. Также предварительное инфицирование слабопатогенным штаммом ВИЧ-2 защитило одну из шести макак от дальнейшего суперинфицирования патогенным штаммом SIVmac25l при введении вируса в прямую кишку.
Развитие защиты коррелировало с высокой концентрацией антител и перекрестно реагирующих нейтрализующих антител. Однако предотвращение суперинфицирования одного животного не позволило получить окончательной информации о том, какие иммунные реакции обеспечивали такую защиту. Аналогичное исследование показало возможность развития защиты от инфицирования SIV после первичного заражения ВИЧ-2. Предварительное инфицирование одним штаммом ВИЧ-2 также защищало бабуинов от суперинфицирования гетерологичным штаммом ВИЧ-2.
Напротив, предварительное инфицирование непатогенным SHIV не защищало от инфицирования патогенным SIV. В последнем случае, однако, могло не происходить достаточной репликации SHIV.
В нескольких исследованиях было сделано общее наблюдение, свидетельствующее о существовании определенного порога напряженности репликации вакцинного ослабленного, но инфекционного вируса, достижение которого необходимо для индукции защитных иммунных реакций. Эти данные о существовании «порогового эффекта» могут способствовать выявлению коррелятов развития противовирусной защиты в результате вакцинации.
В другом варианте при введении ослабленного мутантного вируса с делецией в гене nef, не вызывающего развития заболевания у макак резус, происходит выработка высокого титра антител против вируса. Активация клеточных иммунных реакций изучена не была. Последующее введение таким животным вируса дикого типа в количестве до 1000 инфекционных доз для макак резус приводило к развитию защиты от инфицирования.
Аналогичные данные позднее были получены другими группами исследователей при использовании SIV и SHIV с делецией в гене nef. В одном случае развитие защиты не коррелировало с выработкой нейтрализующих антител. В другом была продемонстрирована важная роль ЦТЛ, активных в отношении Env. Описана корреляция развития защиты от введения патогенного SIV с цитотоксическими и нецитотоксическими реакциями CD8+-клеток; последние зависели от нехемокиновых противовирусных растворимых факторов. В одном интересном исследовании показано развитие защиты от введения инфицированных вирусом клеток.
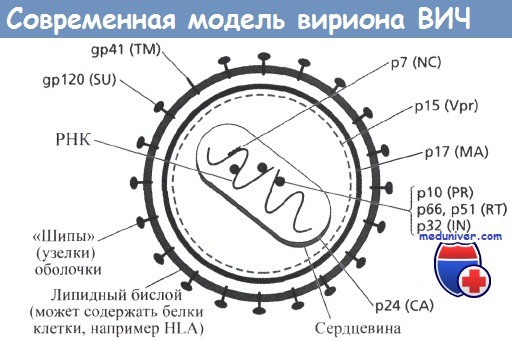
Точная локализация белков Nef и Vif, связанных с сердцевиной, до сих пор не установлена.
Использованы рекомендованные сокращения названий вирусных белков.
В других исследованиях у макак резус, ранее инфицированных штаммом SIV, ослабленным по nef, впоследствии развивалась устойчивость к воздействию штамма SHIV. Поскольку область генома, кодирующая белки оболочки вирусов у двух таких штаммов различалась, развитие защиты должно быть связано с клеточноопосредованной иммунной активностью, а не с выработкой нейтрализующих антител. Во всех этих исследованиях было показано улучшение экспрессии вирусных антигенов в отсутствие белка Nef. Этот белок вируса может угнетать экспрессию инфицированными клетками комплексов вирусных антигенов с молекулами главного комплекса гистосовместимости (МНС) I класса. За счет этого происходит снижение противовирусной иммунной активности CD8+-клеток организма-хозяина.
В других исследованиях были изучены инфекционные SIV, у которых отсутствовали гены nef vpr и последовательности нуклеотидов U3, расположенные «выше по течению», а также SIV, у которого отсутствовал ген nef. Развитие защиты было связано с высокой активностью репликации вакцинных штаммов вирусов в организме животного. Отмечалась некоторая корреляция с содержанием нейтрализующих и просто связывающихся противовирусных антител. Кроме того, использование ослабленного штамма SIV позволило защитить трех обезьян от инокуляции высокой дозы инфекционного SHIV.
При этом не наблюдалось как предварительного образования нейтрализующих антител против SHIV, так и их появления в течение года наблюдений после инокуляции SHIV Развитие защиты от абсолютно гетерогенной оболочки должно быть связано с клеточноопосредованными реакциями и, возможно, реализуется с участием системы врожденного иммунитета.
Другие исследования SIV с различными независимыми или комбинированными мутациями (например, генов vpr, vpx и vif) дали сходные результаты. Во многих работах была показана корреляция между развитием защиты и репликацией ослабленного вируса до высоких концентраций вскоре после вакцинации. Как показывает пороговая гипотеза, только ослабленные вирусы, вызывающие развитие пролонгированной, достаточно сильной вирусемии, обеспечивают резистентность организма к введению SIV дикого типа. Должно установиться состояние защитного иммунитета.
Кроме того, у макак резус, последовательно иммунизированных SHIV с делецией по генам vpu и nef а затем одним мутантным SIV с делецией по гену vpu, происходила выработка нейтрализующих антител и активация ЦТЛ, которые распознавали как гомологичный, так и гетероло-гичный SHIV и SIV. При введении вируса через слизистую оболочку прямой кишки происходило инфицирование всех вакцинированных животных, но уровень вирусемии в плазме был снижен в два - пять раз, и у вакцинированных животных поддерживалась стабильная концентрация CD4+-Т-клеток. По-видимому, происходило развитие защиты от заболевания, по крайней мере на короткий промежуток времени.
Способность живого ослабленного SIV защищать от развития заболевания может быть связана с первичной индукцией выраженных реакций 1 типа и выработкой b-хемокинов. В связи с этим введение живого ослабленного SIV совместно с патогенным вирусом не защищало от развития заболевания. Очевидно, что для защиты от вирулентного вируса существует необходимость предварительного формирования состояния иммунокомпетентности.
Возможные проблемы при работе с ослабленными вирусами в вакцинах от ВИЧ
Использование живых ослабленных вирусов сопряжено с определенными трудностями, которые связаны с тем, что инокуляция некоторых таких вирусов (например, с делецией по гену nef) новорожденным макакам может вызывать развитие СПИДа. Кроме того, у некоторых взрослых животных, которым вводили такие ослабленные вирусы, через несколько месяцев или лет после воздействия вируса развивалось заболевание.
Иногда у животных, которым был введен ослабленный вирус, развилось заболевание после репарации делетированного гена in vivo. Необходимо учитывать такой риск реверсии мутантного вакцинного вируса, несущего делецию заданных генов, в вирус дикого типа. Введение мутантов SIV с делецией по nef также не всегда предотвращало инфицирование гетерологичными штаммами вируса.
Проблема использования ослабленных вирусов может быть решена за счет использования условно живых разновидностей ВИЧ, репликация которых может быть индуцирована с помощью временного введения лекарственных препаратов, например доксициклина. Отмена препарата приводит к прекращению репликации вируса. Однако интеграция ослабленного вируса может привести к развитию других патологических процессов (например, опухолевого роста). Таким образом, использование такого подхода в случае, если не будет разработано других эффективных стратегий с использованием неинфекционных вирусов, других вирусных векторов или белков ВИЧ, будет связано с определенными трудностями.
Тем не менее при использовании в качестве модели ослабленный вирус обладает наиболее близким сходством с относительно стабильно реплицирующимся вирионом, инфицирующим организм хозяина.
В этом случае возможно поддержание эффективного противовирусного ответа и контроля над патогенной активностью вируса. Таким образом, продолжается проведение исследований с использованием ослабленных вирусов, которые могут давать информацию, связанную с иммунными реакциями, необходимыми для развития защитного иммунитета против ВИЧ.
- Читать "Эффективность белков оболочки ВИЧ в вакцине"
Оглавление темы "Вакцины (прививки) от ВИЧ":- Свойства идеальной вакцины от ВИЧ
- Эффективность цельных инактивированных вирусов в вакцинах от ВИЧ
- Эффективность цельных ослабленных вирусов в вакцине от ВИЧ
- Эффективность белков оболочки ВИЧ в вакцине
- Эффективность белка Tat ВИЧ в вакцине
- Эффективность векторов белков ВИЧ в осповирусах
- Эффективность векторов белков ВИЧ в аденовирусах
- Эффективность векторов белков ВИЧ в редких вирусах
- Эффективность вакцины от ВИЧ на основе сердцевины вируса
- Эффективность ДНК-вакцин от ВИЧ