Влияние белков ВИЧ на клетку
Вирусный белок Tat вместе с клеточными РНК-связывающими белками (возможно, фосфорилированными) воздействует на последовательности нуклеотидов TAR в вирусной LTR.
Tat - transactivatore of transcription - вирусный белок, усиливающий транскрипцию с вирусных генов, в том числе из положения «транс», в десятки раз.
Экспрессия вируса значительно увеличивается после взаимодействия клеточных белков с другими областями LTR. Клеточные белки, влияющие на активность Tat, способствуют связыванию Tat с TAR-элементом LTR или с такими промоторами, как АР-1, расположенными впереди TAR.
Таким образом, два типа взаимодействий Tat с вирусной LTR могут приводить к усилению репликации ВИЧ. Индукция выработки TNF Т-клетками под действием Tat ВИЧ также может способствовать увеличению выработки вирусных частиц.
Другие ранние исследования показали, что два родственных Tat-связывающих белка могут конкурировать за усиление (например, MSSI) или ослабление (например, ТВР-1) активности Tat и таким образом влиять на продукцию ВИЧ.
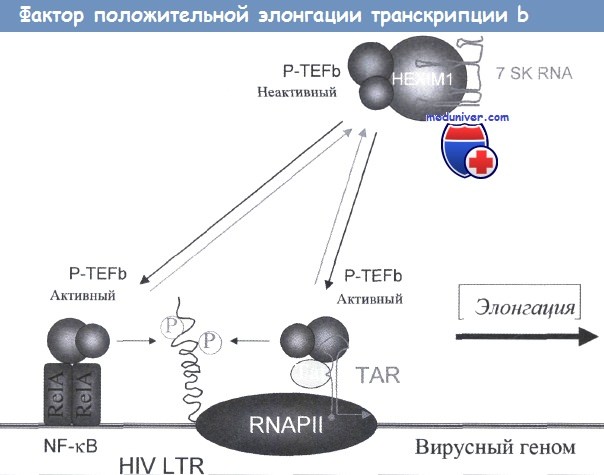
В этом отношении такие клеточные факторы, как циклин Т1, гексим (hexim) и 7SK РНК, могут влиять на степень активности Tat путем активации или инактивации фактора позитивной элонгации транскрипции b (P-TEFb), поддерживающего элонгацию транскрипции мРНК.
Этот комплекс вместе с Tat способствует транскрипции РНК и с вирусных LTR. Белок Nef также может влиять на репликацию вируса за счет взаимодействия с клеточными факторами, которые участвуют в активации клетки.
Вирусный промотер содержит большое количество обычных промотерных элементов, а также энхансерных последовательностей и трансактивационных (TAR) элементов.
Они привлекают РНК-полимеразу II (RNAPII) и устанавливают на длинном концевом повторе (LTR) ВИЧ. Затем RNAPII проходит промотер и останавливается в TAR-элементе или возле него.
P-TEFb, состоящий из циклина Tl (CycTl) и циклин-зависимой киназы-9 (CDK9), необходим для элонгации транскрипционных комплексов. Он фосфорилирует С-концевой домен RNAPII.
После этого происходит отделение факторов инициации и отрицательной транскрипции от RNAPII и замещение их молекулярными механизмами кэппирования, сплайсинга и полиаденилирования.
P-TEFb существует в клетке в двух формах. Более крупный, неактивный комплекс, содержит CycTl, неактивную CDK9-киназу, HEXIM1 и 7SK РНК.
Активный комплекс меньшего размера состоит только из CycTl и CDK9 и необходим для действия NF-kB и Tat. Большинство клеток содержат достаточные количества активного P-TEFb для обеспечения репликации ВИЧ.
Связывание NF-kB с P-TEFb приводит к инициации первого цикла вирусной транскрипции.
Однако после образования достаточного количества Tat этот вирусный белок и P-TEFb связываются с TAR в LTR с высокой аффинностью, что приводит к существенному увеличению скорости репликации вируса.
Важно отметить, что как NF-kB, так и Tat обеспечивают правильную транскрипцию и процессинг вирусных генов.
- Вернуться в оглавление раздела "Инфекционные болезни"
Оглавление темы "Репликация ВИЧ":- Этапы репликации ВИЧ внутри клетки
- Влияние внутриклеточных факторов на репликацию ВИЧ
- Особенности репликации ВИЧ в Т-клетках
- Особенности репликации ВИЧ в моноцитах и макрофагах
- Белок APOBEC3G как механизм резистентности к ВИЧ
- Аналоги белка APOBEC3G у животных
- Природные функции белков APOBEC
- Белок TRIM5a как механизм резистентности к ВИЧ
- Влияние цитокинов на репликацию ВИЧ
- Влияние белков ВИЧ на клетку