Нервная регуляция длины мышц. Иннервация мышц
Как было показано ранее, рефлекторные дуги моносинаптических и полисинаптических рефлексов способствуют достижению различных целей. Полисинаптические рефлекторные дуги обеспечивают защитные рефлексы (рефлексы отступления), в то время как моносинаптические рефлексы включены в функциональные рефлекторные кольца обратной связи, регулирующие длину и тонус скелетных мышц. На самом деле, каждую мышцу контролируют две системы обратной связи:
- система контроля длины мышцы, в которой рецепторами, определяющими изменение длины мышцы, служат волокна с ядерной сумкой в мышечных веретенах;
- система контроля тонуса, в которой рецепторами, определяющими изменение тонуса, служат сухожильный орган Гольджи и волокна с ядерной цепочкой в мышечных веретенах.
Рецепторы растяжения и напряжения. Мышечные веретена служат как рецепторами растяжения (удлинение) мышцы, так и рецепторами ее напряжения. Каждому из этих видов чувствительности служат отдельные интрафузальные волокна: волокна с ядерной сумкой и волокна с ядерной цепочкой соответственно. Оба вида интрафузальных волокон обычно короче и тоньше экстрафузальных мышечных волокон. Для наглядности эти два вида интрафузальных волокон на рисунке изображены раздельно, хотя на самом деле более короткие и тонкие волокна с ядерной цепочкой прилежат непосредственно к волокнам с ядерной сумкой, которые чуть длиннее первых.
Два волокна с ядерной сумкой и 4—5 волокон с ядерной цепочкой, как правило, составляют одно мышечное веретено. Посередине волокно с ядерной сумкой раздается вширь так, что получается сумка, вмещающая около 50 ядер. Снаружи сумку обвивает сеть из чувствительных нервных волокон, которые называют первичными окончаниями (раньше их называли аннулоспиральными). Эти окончания чрезвычайно чувствительны к растяжению мышцы и регистрируют главным образом изменение мышечной длины. Таким образом, волокна с ядерной сумкой являются рецепторами растяжения. Волокна с ядерной цепочкой, напротив, регистрируют преимущественно состояние постоянного растяжения мышцы и, следовательно, являются рецепторами напряжения.
Поддержание постоянной длины мышц. Экстрафузальные мышечные волокна в состоянии покоя имеют определенную длину, которая всегда должна поддерживаться организмом неизменной. Растяжение мышцы сверх этой длины влечет за собой и растяжение мышечного веретена. При этом в первичных окончаниях возникают ПД, которые со скоростью до 120 м/с устремляются по афферентным волокнам группы IA в спинной мозг, где через синапсы передаются на мотонейроны передних рогов спинного мозга.
Те, в свою очередь, возбуждаются и разряжаются нервными импульсами, которые так же стремительно мчатся по толстым быстропроводящим а1-эфферентным волокнам назад, к рабочим экстрафузальным мышечным волокнам, заставляя их сократиться до прежней длины. Такую реакцию вызывает любое растяжение мышцы.
Сохранность этого рефлекторного кольца врачи проверяют быстрым и коротким ударом неврологического молоточка по сухожилию мышцы, например, при исследовании коленного рефлекса — по четырехглавой мышце бедра, вследствие чего мышца растягивается и запускает моносинаптический рефлекс.
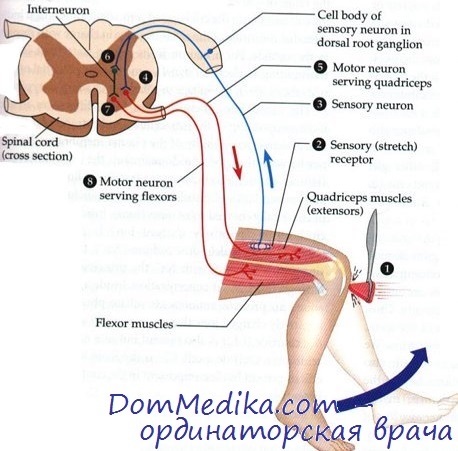
Поскольку рефлекторная дуга для отдельной мышцы захватывает лишь 1 или 2 нервных корешка (сегментов спинного мозга), то выявление патологического рефлекса позволяет врачу довольно точно предположить соответствующий уровень поражения корешков или спинного мозга. Таким образом, в клинической неврологии исследование глубоких мышечных рефлексов имеет немаловажное значение для поиска уровня поражения. Наиболее важные глубокие мышечные рефлексы, сегменты, на уровне которых замыкаются их рефлекторные дуги, и способы их вызывания представлены на рисунке. Однако не следует забывать, что клиническое исследование глубоких мышечных рефлексов на практике — это сугубо экспериментальный процесс, поскольку такое кратковременное растяжение мышцы, как следующее за ударом неврологического молоточка, в повседневной жизни практически не встречается.
Рефлекторное расслабление мышц-антагонистов. Рефлекторное сокращение растянутой мышцы, возвращающее ее длину, сопровождается рефлекторным расслаблением мышцы-антагониста (или группы мышц-антагонистов). Дуга описанного рефлекса, вероятно, берет свое начало в мышечных веретенах. В составе бессчетных мышечных веретен волокна с ядерной цепочкой, кроме первичных окончаний, о которых говорилось выше, содержат вторичные окончания, именуемые также гроздевидными (у-кустовидные) нервными окончаниями.
Вторичные окончания реагируют на растяжение мышцы, так же как и первичные, с тем лишь различием, что возникающая в них импульсация устремляется в ЦНС не по таким же волокнам, как идущие от первичных окончаний волокна группы IA, а по более тонким волокнам группы II. Далее нервные импульсы передаются через вставочные нейроны спинного мозга и вызывают торможение в сети, приводящее к расслаблению мышц-антагонистов (реципрокное антагонистическое торможение).
Регулировка заданной длины мышцы. Существует специальная двигательная система, функция которой заключается в регулировке длины мышцы путем влияния на рефлекторную дугу.
Передние рога спинного мозга содержат не только крупные а-мотонейроны, но и мелкие у-мотонейроны. Их аксоны, именуемые y-волокнами, оканчиваются на мелких поперечнополосатых интрафузальных волокнах в мышечных веретенах. Возбуждение, спускаясь по у-волокнам, заставляет сокращаться интрафузальные мышечные волокна на том и другом концах мышечного веретена. Те, сокращаясь, растягивают середину мышечного веретена — каждый в свою сторону, из-за чего в первичных нервных окончаниях возникают ПД, которые, в свою очередь, повышают тонус рабочей мышцы.
На у-мотонейроны влияют несколько нисходящих двигательных путей, включая пирамидный, ретикулоспинномозговой и преддверно-спинномозговой. Таким образом, у-мотонейроны оказываются посредником в системе контроля мышечного тонуса, осуществляемого высшими двигательными центрами, без сомнения, играющими важную роль в регуляции произвольных движений. у-Эфферентные волокна позволяют точно регулировать произвольные движения и, кроме того, — чувствительность рецепторов растяжения. Сокращение интрафузальных мышечных волокон, влекущее за собой растяжение средней части мышечного веретенa, снижает порог возбуждения рецепторов растяжения, то есть для их активации потребуется совсем небольшое растяжение мышцы.
Фузимоторная (у-) иннервация мышцы в норме позволяет автоматически поддерживать заданную длину мышцы.
Медленное растяжение первичных рецепторов (волокна с ядерной сумкой, обвитые аннулоспиральными окончаниями) одновременно с вторичными рецепторами (волокна с ядерной цепочкой с прикрепленными к ним гроздевидными окончаниями) влечет за собой статический, то есть не меняющийся во времени, ответ рецепторов мышечных веретен. Если же первичные рецепторы, напротив, растягиваются очень быстро, возникает динамический (быстро меняющийся) рефлекторный ответ.
Как статический, так и динамический, рефлекторные ответы контролируются эфферентными (двигательными) у-нейронами.
Статические и динамические у-мотонейроны. Считают, что существует два типа у-мотонейронов — динамические и статические. Динамические у-мотонейроны иннервируют преимущественно интрафузальные волокнами с ядерной сумкой, а статические у-мотонейроны, главным образом, — интрафузальные волокна с ядерной цепочкой. Возбуждение, переданное динамическими у-нейронами на волокна с ядерной сумкой, вызывает сильный динамический ответ, опосредуемый первичными окончаниями; в то время как возбуждение, переданное статическими у-нейронами на волокна с ядерной цепочкой, вызывает статический, тонический, ответ.
- Читать "Нервная регуляция мышечного тонуса. Адаптация тонуса мышц"
Оглавление темы "Неврология":- Моносинаптические и полисинаптические рефлексы. Рефлекторная дуга
- Нервная регуляция длины мышц. Иннервация мышц
- Нервная регуляция мышечного тонуса. Адаптация тонуса мышц
- Задний и передний спиномозжечковые пути
- Задние канатики спинного мозга. Признаки поражения задних канатиков
- Передний спиноталамический путь: анатомия, признаки поражения
- Латеральный спиноталамический путь: анатомия, признаки поражения
- Обработка соматосенсорной информации в ЦНС. Стереогноз и астереогноз
- Причины нарушения чувствительности. Патология соматосенсорных путей
- Двигательные зоны коры коры головного мозга