Онтогенез В-лимфоцитов - дифференциация
Количество специфичностей В-клеток и продуцируемых ими иммуноглобулинов чрезвычайно велико. Однако в зародышевой клетке не предсуществуют гены антител к каждому изолированно взятому антигену. Генерация репертуара антител во многом обусловлена соматическими комбинаторными механизмами на уровне генных сегментов ДНК. Молекула иммуноглобулина состоит из двух легких (L) и двух тяжелых (Н) полипептидных цепей. Существует 2 типа L-цепей (к и X) и 5 классов Н-цепей (u, у, а, q, s), a также их подтипы и подклассы. В молекуле иммуноглобулина две идентичные L-цепи и две идентичные Н-цепи.
Строение L- и Н-цепей иммуноглобулина однотипное: они состоят из вариабельной (V) и константной (С) области, различия касаются числа доменов в С-области (в L-цепях их по одному, в Н-цепях по 3—4). Тип, или класс, иммуноглобулина обусловлен С-областями Н- и L-цепи, а его специфичность — V-областью. При объединении V-областей Н- и L-цепи в молекуле иммуноглобулина образуется два специфических антигенсвязы-вающих участка.
Н- и L-полипептидные цепи иммуноглобулина не наследуются как нечто целое. Напротив, генные сегменты, кодирующие различные участки этих белков, в молекуле ДНК разобщены. Они начинают группироваться, объединяться и приобретать новую конфигурацию (перестройка, или реаранжировка, генов иммуноглобулина) только после того, как лимфоцит избирает В-линейную направленность дифференцировки. Ген V-области L-цепей собирается из 2 сегментов (V и J), а ген V-области Н-цепей — из 3 сегментов (V, D и J): V — вариабельный, D — разнородный, J — объединяющий.
Полный ген иммуноглобулиновых цепей включает наряду с перечисленными сегментами участок ДНК для С-области L-цепи (к, А.) и Н-цепи (ц, 5, у, а, е). В литературе приводятся данные о разном количестве генов для каждого из сегментов (V, D, J): так, например, для V-генов Н-цепей от 50-59 до 500 - 1000. Количество генов V-сегментов, как правило, преобладает. Так, число D-генов Н-цепей иммуноглобулина составляет 15—30, а J-генов — 4—6. В ходе созревания В-клетки в ней используется лишь по одному из существующих V-, J- и D-генов, что позволяет образовывать в результате комбинаций и сочетания VH и VL огромное разнообразие антительных специфичностей.
Процесс реаранжировки генов иммуноглобулина контролируется комплексом ферментов, носящих общее название «V(D)J-рекомбиназа». Особенностью данного комплекса ферментов является то, что в его состав входит гетеродимерная эндонуклеаза RAG-1/RAG-2, кодируемая генами, активирующими рекомбинацию (recombination-activating genes), необходимая на начальных этапах расщепления ДНК в ходе реаранжировки генов иммуноглобулина. Другим специфическим компонентом V(D)J-эндонуклеазного комплекса является TdT — фермент, функционирующий в течение очень короткого периода дифференцировки В-клеток (только на этапе сборки генов Ig-цепей).
TdT обеспечивает нематричный синтез N-вставок нуклеотидных последовательностей в местах разрывов ДНК, возникающих в процессе объединения V-, D- и J-сегментов. Образующаяся в результате реаранжировки последовательность ДНК выглядит следующим образом: V-N-D-N-J. N-вставки обеспечивают дополнительную вариабельность иммуноглобулинов. TdT является ядерным маркером ранних костно-мозговых этапов дифференцировки В-клеток.
Таким образом, формированию зрелых В-лимфоцитов, имеющих функционально активный BCR, предшествует этап антигеннезависимой дифференцировки В-линейных предшественников (В-ЛП). Этот этап характеризуется не только реаранжировкой генов иммуноглобулина, но также изменениями иммунофенотипа клеток. Последовательность изменений в ходе антигеннезависимой (костно-мозговой) дифференцировки В-лимфоцитов представлена в таблице. Созревание В-лимфоцитов в костном мозге может быть разделено на 3 стадии на основании экспрессии молекул иммуноглобулинов. На наиболее ранних стадиях созревания В-клеток (про-В) такие молекулы не экспрессируются. Про-В-клетки идентифицируются на основании В-линейных антигенов, белков комплекса BCR и реаранжировки иммуноглобулиновых генов.
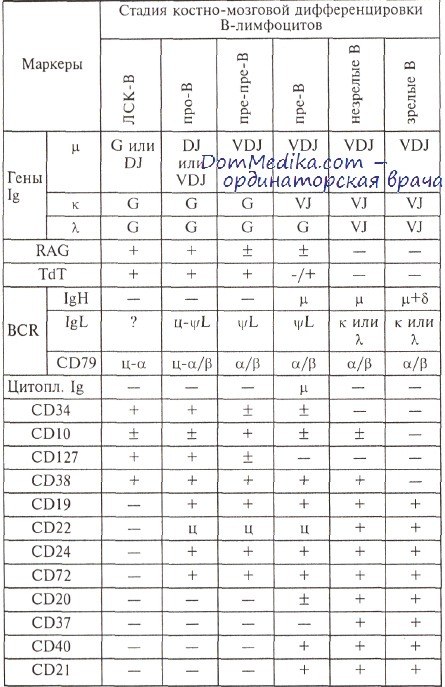
В-ЛП следующего этапа дифференцировки (пре-В) не имеют мембранных иммуноглобулинов, но содержат в цитоплазме Н-цепь IgM (ц-цепь). Между этими этапами существует промежуточная стадия пре-пре-В, которую отличают не все авторы. Клетки этого этапа дифференцировки являются нормальными аналогами самого частого варианта острых лимфобластных лейкозов и лимфом из В-линейных предшественников — так называемого общего (от англ. common) варианта. Сложность его идентификации в нормальной дифференцировке обусловлена отсутствием полной ясности в последовательности появления CD19 и CD10 на нормальных В-лимфоидных предшественниках. Еще одна стадия дифференцировки незрелых В-предшественников — это транзиторные В-клетки, т. е. пре-В-клетки, экспрессирующие на мембране ц-цепь в комплексе с суррогатными Х-цепями.
Наиболее зрелый фенотип В-ЛП характеризуется наличием мембранных иммуноглобулинов (М или M+D, незрелые и зрелые В-клетки соответственно) одного из типов L-цепей — к или X.
Первые клетки В-линии обнаруживаются в печени и сальнике плода человека с 8-й недели внутриутробного развития. В период 8—14 нед В-лимфоцитопоэз продолжается в печени и перемещается в костный мозг. С 4-го месяца после рождения костный мозг становится основным, а затем единственным органом В-лимфоцитопоэза. Зрелые sIg+ В-лимфоциты обнаруживаются в селезенке и лимфатических узлах плода с 14-й недели.
Процесс дифференцировки В-линейных предшественников в костном мозге — это по существу формирование функционально зрелого В-рецепторного комплекса на мембране В-лимфоцитов. Для этого происходят реаранжировка генов иммуноглобулинов, экспрессия молекул IgM, CD79 (Iga и Igb) и В-линейно-ассоциированных молекул на мембране клеток.
- Читать "Перестройка иммунноглобулинов при В-лимфоцитопоэзе"
Оглавление темы "Гемопоэз":- Механизмы миграции стволовых кроветворных клеток (СКК)
- Мезенхимная стволовая клетка - свойства, функции
- Эмбриональная стволовая клетка (ЭСК) - свойства, функции
- Пластичность стволовых клеток - возможности дифференцировки
- Врожденный и адаптивный иммунитет - механизмы
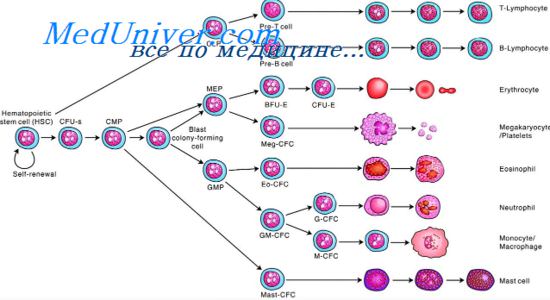
- Стволовые лимфоидные клетки - общий лимфоидный предшественник
- Онтогенез В-лимфоцитов - дифференциация
- Перестройка иммунноглобулинов при В-лимфоцитопоэзе
- Антигены и рецепторы В-лимфоцитов
- Созревание В-лимфоцитов в костном мозге