Механизмы эритропоэза и его регуляция
В регуляции эритропоэза решающую роль играет эритропоэтин, в отсутствие которого БОЕ-Э не пролиферируют либо делятся очень медленно. Синтез гемоглобина в эритробластах также ингибирован. Очевидно, что от уровня эритропоэтина зависит интенсивность эритропоэза. Основное место продукции этого цитокина — почки, здесь его продукция регулируется напряжением кислорода. Во всех случаях, когда рО2 снижается (кровопотеря, подъем на высоту, заболевания легких и др.), происходит стимуляция эритропоэза.
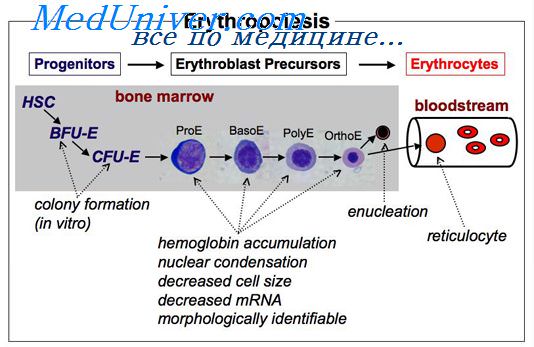
При заболеваниях почек часто развивается анемия за счет снижения/прекращения выработки эритропоэтина, что подтверждается его выраженным терапевтическим эффектом при такой патологии. Эритропоэз в конечном счете представляет собой многоступенчатый процесс дифференцировки БОЕ-Э и КОЕ-Э через стадии морфологически распознаваемых ранних и поздних эритробластов и ретикулоцитов в зрелые эритроциты. В течение этого процесса происходит последовательное образование проэритробластов, базофильных, полихроматофильных и оксифильных эритробластов, позитивно регулирующееся эритропоэтином.
На разных стадиях эритроидной дифференцировки влияние эритропоэтина неодинаково: проэритробласты отвечают на него усилением пролиферации, созревания и предотвращением апоптоза. Экспрессия антиапоптических генов в этих клетках очень низкая, и незрелые эритробласты практически полностью уязвимы для индукторов этого процесса в отсутствие эритропоэтина. Последний подавляет апоптоз посредством индукции экспрессии гена Bcl-XL, члена семейства Bcl-2, вовлеченного в защиту от программированной клеточной гибели.
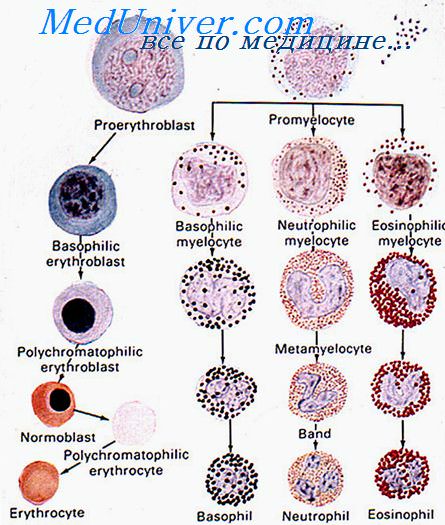
Экспрессия Bcl-XL очень низка во время ранней эритроидной дифференцировки и увеличивается в средних и поздних эритробластах одновременно с потерей зависимости от эритропоэтина. Таким образом, уровень эритропоэтина детерминирует судьбу клеток эритроидной дифференцировки путем поддержания выживания ранних эритробластов.
В костном мозге эритропоэз происходит в дискретных анатомических единицах — эритробластных островках, состоящих из одного или двух макрофагов-кормушек, окруженных одним или несколькими кольцами из эритробластов разной стадии созревания. Внутренний слой кольца эритробластов содержит незрелые клетки, а более зрелые находятся на периферии островка. Пространственное распределение зрелых и незрелых эритробластов может играть важную роль в эритропоэзе, так как межклеточные взаимодействия необходимы для нормального функционирования эритрона.
Созревающие эритробласты могут передавать ингибирующий сигнал соседям, так как их созревание снижает потребность организма в эритроидных клетках. Экспрессия Fas (рецептор, активация которого индуцирует апоптоз) на эритробластах повышается при снижении уровня эритропоэтина. Интересно, что концентрация эритропоэтина, необходимая для предотвращения Fas-индуцированного апоптоза, сравнима с таковой в сыворотке крови больных с низким гемоглобином или сниженной массой эритроцитов.
Экспрессия Fas по мере созревания ранних эритробластов усиливается и сохраняется на высоком уровне на стадиях терминальной дифференцировки. Fas-связывание эффективно только у менее зрелых эритробластов, практически на базофильном уровне, если индукция апоптоза необходима как противовес высокого уровня эритропоэтина. Напротив, Fas-лиганд селективно экспрессирован в поздних дифференцированных Fas-чувствительных эритробластах, в основном на оксифильной стадии. Fas-лиганд функционален в зрелых эритробластах и может вызывать гибель Fas-чувствительных эритробластных мишеней Fas-зависимым апоптозом. Важно, что несущие Fas-лиганд зрелые эритробласты могут вызывать Fas-зависимую клеточную гибель незрелых эритробластов. Высокие дозы эритропоэтина аннулируют данный эффект. Это свидетельствует о наличии негативной обратной регуляции в эритробластном островке.
Зрелые, но не терминально дифференцированные клетки оказывают цитотоксический эффект на свои ближайшие предшественники. Гипотетически эта негативная регуляция связана с низким уровнем эритропоэтина. Она ингибируется при повышении его продукции при стимуляции эритропоэза. Взаимодействие Fas и Fas-лиганда может представлять механизм апоптозопосредованного контроля эритропоэза.
Важно отметить, что в регуляции эритроидной дифференцировки число новообразованных зрелых клеток (эритроцитов) не играет решающей роли для интенсивности эритропоэза: даже при выраженной полицитемии эритропоэз усиливается дозозависимым образом при введении экзогенного эритропоэтина. Эффект совершенно независим от степени полицитемии и числа эритроцитов в циркуляции. Регуляция определяется в основном физиологической функцией зрелых клеток — переносом кислорода, его напряжением в почках.
- Читать "Лейкопоэз (гранулоцитопоэз) - этапы, механизмы, регуляция"
Оглавление темы "Гемопоэз":- История изучения кроветворения - гемопоэза
- Стволовая кроветворная клетка (СКК) - история изучения, свойства
- Дифференциация стволовых кроветворных клеток - механизмы
- Механизмы эритропоэза и его регуляция
- Лейкопоэз (гранулоцитопоэз) - этапы, механизмы, регуляция
- Лимфоцитопоэз - этапы, механизмы, регуляция
- Факторы транскрипции гемопоэза - транскрипционный контроль
- Tal-1/SCL - функции фактора транскрипции гемопоэза
- Rbtn2/LMО2 - функции фактора транскрипции гемопоэза
- GATA1, GATA2 и GATA3 - функции факторов транскрипции гемопоэза