Иммунитет при скрепи - механизмы защиты
Одной из наиболее интересных загадок скрепи (как, впрочем, и других представителей группы губкообразных энцефалопатий) является отсутствие клеточной и гуморальной иммунологической реакции хозяина па инфекционный агент.
Chandler использовал 10 разнообразных серологических приемов, но не добился успеха в попытках определить антитела к агенту скрепи в сыворотках овец или коз. Антитела не удалось обнаружить ни в реакциях нейтрализации и связывания комплемента, ни в прямой и в непрямой модификациях МФА (Gibbs е. а., А. М. Гардашьяп и др., Porter е. а.). Отрицательные результаты были получены и при исследовании животных в инкубационном периоде, и в разгаре заболевания, и даже после гипериммунизации животных на протяжении более 5 мес (А. М. Гардашьяп и др., Gardiner, Eklunde. а.).
Сходный результат был получен Albrecht, пытавшимся иммунизировать мышей инфекционным и инактивированным препаратами мозговой ткани с адъювантом Фрейнда. Этот же исследователь установил, что лимфоциты селезенки не трансформируются в присутствии нормального и инфицированного агентом скрепи мозга, ио при этом сохраняют способность трансформироваться под влиянием фитогемагглютинипа.
Не удалось продемонстрировать наличие в тканях иммунного комплекса при скрепи с помощью метода флуоресцентного окрашивания с анти-IgG или антикомплементом (С3); не удались попытки снизить инфекционный титр агента иммунной преципитацией с антисывороткой, которая, как ожидалось, содержит антитела к агенту скрепи; наконец, неполучены доказательства присутствия в кровяном русле комплекса инфекционный агент— антитело (Gibbs, Gajdusek).
Ни в инкубационном периоде, ни во время клинически выраженного заболевания не было обнаружено интерферона в тканях инфицированных скрепи животных. В то же время зараженные скрепи мыши сохраняли способность индуцировать синтез иптерферона в ответ на заражение вирусами везикулярного стоматита или Западного Нила (Katz, Koprowski).
Не дали положительных результатов и разнообразные попытки стимулировать интерферон в организме инфицированных агентом скрепи животных с тем, чтобы таким образом повлиять па течение инфекционного процесса. Так, например, еженедельное введение статолона в течение 7 мес не оказало влияния на течение инфекционного процесса у мышей (Field е. а.), равно как и введение поли-И: поли-Ц не влияло на особенности клинического течения и смертность животных (Worthington).
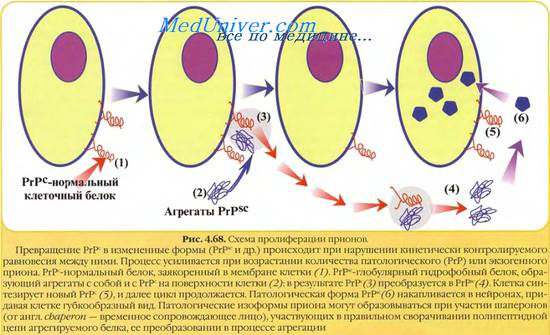
В последние несколько лет появилась серия сообщений, посвященных исследованию форменных элементов белой крови при скрепи. В результате было показано, что при внутримозговом заражении мышей возбудителем скрепи клиника заболевания развивается через 4 мес, но уже с 3-го дня наблюдается уменьшение числа нейтрофилов в крови. Фактор, понижающий количество нейтрофилов, обнаруживался вместе с возбудителем скрепи в мозге и селезенке больных овец и мышей. Титр этого фактора в мозговой ткани зараженных мышей составлял около 1012, а титр возбудителя скрепи — около 105.
Пассажами на мышах сильно разведенного инфекционного материала (10-8—10-9) удалось изолировать фактор, понижающий количество нейтрофилов. Оказалось, что введение его мышам в концентрации 1012 единиц не приводит к развитию скрепи. Следовательно, агент скрепи и фактор, вызывающий снижение числа нейтрофилов при этом заболевапии, не являются идентичными, хотя, как показали специальные исследования, фактор проходит фильтры с размерами пор 220, 100 и 50 нм, задерживается фильтром с размером пор 25 им (Licursi е. а., Carp е. а.).
Интересные данные получены и при несколько ином аспекте изучения форменпых элементов белой крови. Field и Shenton (1973) исследовали возможность иммунологического дифференцирования агента скрепи от нормальных антигенов селезенки и мозга мышей чувствительных линий. Для этого суспензии инфицированного мозга инокулировали морским свинкам, а в качестве контрольного антигена использовали гомогенаты нормального мозга мышей или гомогенаты мозга мышей с искусственно индуцированной астроцитарной гипертрофией. О наличии иммунитета судили по высвобождению из иммунных лимфоцитов морских свинок фактора, угнетающего миграцию макрофагов.
На 5—12-й день после иммунизации выявили значительные различия в высвобождении такого фактора, угнетающего миграцию макрофагов, при использовании в качестве иммуногепа и тест-антигена гомогената мозга мышей, зараженных скрепи. Такой же результат получен с гомогенатами поражеппой селезенки. Существенно, что подобный же эффект наблюдали и при иммунизации мозгом мышей, страдающих искусственно индуцированной астроцитарной гипертрофией. Авторы полагают, что в основе антигенной альтерации мозга при скрепи лежит поражение астроцитов, и надеются использовать описанный выше прием для специфической и количественной идентификации скрепи, так как выраженность обнаруженного ими эффекта точно коррелирует с биологическими титрами агента.
Эти же исследователи в тесте электрофоретической подвижности макрофагов показали большее повышение чувствительности лимфоцитов морской свинки после инъекции мозговой или селезеночной ткани мышей, зараженных скрепи, по сравнению с нормальной тканью. Эти результаты могут указывать на присутствие в инфицированных тканях антигена агента скрепи, причем различия в реакциях с нормальной тканью и с тканью, содержащей агент скрепи (скрепи — нормальное различие— СНР), можно выражать количественно. Специальные исследования, проведенные на людях в возрасте от 20 лет до 91 года, показали, что СНР увеличивается с возрастом во всех тканях (Field, Shenton) и появляется рапыпе в тканях тимэктомированных животных, что указывает на некую акселерацию процесса повзросления (Field, Shenton).
На основании результатов этих исследований предложен быстрый иммунологический тест на скрепи у овец и коз, для чего сопоставляют реактивность лимфоцитов крови к мозгу (или селезенке) мышей, пораженных скрепи, и реактивность к нормальным тканям, используя тест электрофоретической подвижности макрофагов (Field, Shenton).
С помощью анализа электрофоретической подвижности удалось обнаружить особую белковую фракцию в спинномозговой жидкости больных скрепи животных. Интересно, что эту фракцию пе обнаруживали через 2 и 4 нед после внутрибрюшинного заражения мышей, но обнаруживали у 100% зараженных животных через 6, 8 и 10 нед, она уменьшалась к 12-й неделе и отсутствовала через 14, 16, 18 и 20 нед. Однако во время клинической стадии болезни у 60—70% животных фракция обнаруживалась вновь, что соответствовало 22-й и 24-й педелям от момента заражения. На этом основании и электрофоретический анализ белков спинномозговой жидкости предлагается в качестве лабораторного теста на скрепи у животных (Carp).
- Читать "Эпидемиология скрепи - распространенность"
Оглавление темы "Медленные инфекции":- Иммунитет при скрепи - механизмы защиты
- Эпидемиология скрепи - распространенность
- Возбудитель трансмиссивной энцефалопатии норок
- Клиника трансмиссивной энцефалопатии норок - проявления
- Патогистология трансмиссивной энцефалопатии норок - морфология
- Патогенез трансмиссивной энцефалопатии норок
- Иммунитет при трансмиссивной энцефалопатии норок
- Эпидемиология трансмиссивной энцефалопатии норок - распространенность
- Причины рассеянного склероза - возбудитель
- Клиника рассеянного склероза - проявления