1. Физическое картирование генов. Картирование генов с помощью хромосомных мутаций.
2. Картирование генов с помощью гибридизации in situ.
3. Гибридизация соматических клеток и картирование генов человека.
4. Идентификация генов человека с помощью позиционного клонирования.
5. Создание моделей наследственных болезней. Создание трансгенного животного.
Идентификация генов человека с помощью позиционного клонирования.
Установление локализации гена наследственной болезни в настоящее время можно рассматривать как подготовительный этап для его идентификации, т.е. выделения, клонирования, изучения структуры гена и последовательности нуклео-тидов, а также обнаружения мутаций в этом гене, которые, собственно, можно расценивать как этиологический фактор для соответствующего менделирующего наследственного заболевания.
Классическая молекулярная генетика идентификацию гена обычно начинала с его первичных продуктов — мРНК или белка. Первичные продукты позволяли получить фрагменты гена либо как кДНК, либо как последовательности нуклеотидов, соответствующие последовательности аминокислотных остатков в молекуле белка (такая последовательность может быть короткой, и для ее создания вовсе не надо определять всю последовательность остатков в молекуле соответствующего белка, достаточно определить последовательность 1—2 десятков аминокислот). Эти фрагменты гена превращали в ДНК-зонды, с их помощью идентифицировали мутантный ген в геномных библиотеках или путем гибридизации in situ на препаратах хромосом.
Систематическое изучение генома человека создало предпосылки для реализации иного пути идентификации мутантного гена в тех случаях, когда его первичный продукт неизвестен. Этот путь был назван позиционным клонированием, т.е. поиском гена, исходя из его положения в геноме, которое исходно обычно определяется по результатам генетического картирования. Одним из наиболее ярких примеров идентификации гена с помощью позиционного клонирования является ген муковисцидоза1, хотя он и не является первым идентифицированным таким путем. Ему предшествовало позиционное клонирование гена миодистрофии Дюшенна.
Позиционное клонирование обычно начинают с создания ДНК-зонда на основе полиморфного генетического маркера, тесно сцепленного с геном заболевания. Для гена муковисцидоза было найдено два фланкирующих ген маркера — ген онкогена met и последовательность J3.ll. Однако эти маркеры находились на расстоянии 1600 тыс. п.н., поэтому были предприняты попытки найти более тесно сцепленные с геном полиморфные маркеры. Попытки оказались удачными.
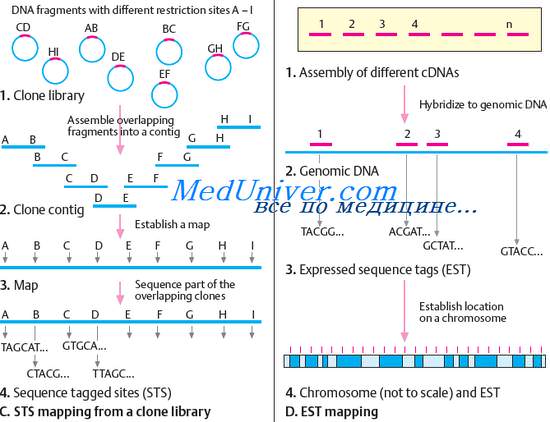
С помощью ДНК-зонда, созданного на основе сцепленного с геном заболевания маркером, из геномной библиотеки извлекают клоны, частично перекрывающиеся по последовательностям с ДНК-зондом, т.е. последний гибридизируется с этими клонами (в работе по картированию гена муковисцидоза использовали большое количество геномных библиотек, как фаговых, так и плазмидных). Теперь ДНК-зонды создают на основе последовательностей нуклеотидов клонов, с их помощью в геномной библиотеке отыскивают перекрывающиеся клоны с новым ДНК-зондом. Вся эта процедура получила название прогулки по хромосоме. Она продолжается до тех пор, пока не будет найдена последовательность ДНК, которая отвечает ряду требований, предъявляемых к искомому гену, о чем будет сказано ниже. Одним из наиболее важных моментов в прогулке по хромосоме является определение правильного направления движения. В работе по клонированию гена муковисцидоза это было сделано с помощью выявления сайтов рестрикции нескольких редко щепящих рест-риктаз в клонах геномной ДНК, которые исследуют по мере прогулки по хромосоме и положение которых в изучаемом сегменте хромосомы установлено заранее.
В настоящее время для выявления перекрывающихся клонов и направления прогулки чаще всего используют так называемые сайты, маркированные сиквенсом (STS). STS представляют собой отсекве-нированные последовательности из нескольких сотен пар нуклеотидов, для которых известны хромосомная локализация и праймеры для ПЦР. Перекрывающиеся клоны легко идентифицируются в ПЦР с одними и теми же праймерами: если в двух клонах из геномной библиотеки одни и те же праймеры обеспечивают ПЦР в этих клонах, значит, они перекрываются. Перекрывающиеся фрагменты ДНК секвениру-ются, и из них строится карта контига, т.е. физическая карта определенной хромосомной области, в которой уже нет перекрывания клонов. На пути к гену муковисцидоза было пройдено 280 тыс. п.н. и велись поиски потенциальных генов. Эти поиски осуществляли путем сравнения последовательностей нуклеотидов исследуемой области с геномной ДНК, полученной от других видов. Предполагали, что структурно сходные последовательности в ДНК других видов будут указывать на то, что они эволюционно консервативны и, следовательно, выполняют важную функцию. Это один из критериев, используемый для поиска генов. Было найдено три консервативные последовательности, из которых от двух как кандидатов в гены муковисцидоза исследователи быстро отказались, а третья консервативная последовательность действительно оказалась началом гена муковисцидоза. В этом удалось убедиться только после того, как было найдено, что третья консервативная последовательность гибридизуется с одной из кДНК, полученной на основе мРНК, выделенных из клеток потовых желез (один из поражаемых органов при муковисци-дозе).
Еще одним критерием обнаружения гена во время прогулки по хромосоме является выявление неметилированных, так называемых ЦГ-островков, характерных для 5'-области экспрессирующихся генов.
Такие ЦГ-островки были найдены в третьей консервативной последовательности, обнаруженной при поиске гена муковисцидоза, как и открытая рамка считывания (еще один критерий полноценного гена). После того как ген муковисцидоза был выделен, оказалось, что он достаточно велик, его размер составил 250 тыс. п.н. и содержит 24 экзона. Теперь надо было получить еще одно доказательство того, что найден «правильный» ген.
Для этого в нем необходимо было найти мутации, которых нет у здоровых индивидуумов. Это было сделано путем сравнения двух последовательностей нуклеотидов клонов, полученных из библиотеки кДНК потовых желез больных муковисцидозом и перекрывающих почти всю кодирующую область гена муковисцидоза с последовательностью нуклеотидов кДНК нормальных индивидуумов. В результате была обнаружена делеция трех пар нуклеотидов в 508-м положении молекулы предсказанного белка. Различными способами подтвердили, что эта мутация не встречается у здоровых людей, зато у больных муковисцидозом она была найдена в 70 % мутантных хромосом. Мутацию назвали AF508.
В настоящее время уже хорошо известно, что это действительно самая частая из более чем тысячи мутаций, обнаруженных в гене муковисцидоза. Белок, кодируемый геном муковисцидоза (ген был назван муковисцидозным трансмембранным регулятором, сокращенно CFTR). Он состоит из 1480 аминокислотных остатков, которые образуют трансмембранные домены и АТФ-связывающие складки. Основная функция CFTR-белка — служить С1_каналом, но возможно, что у него есть еще и другие регуляторные функции. Анализ структуры гена CFTR и самой частой мутации — AF508 позволил вскоре после идентификации гена разработать методы прямой ДНК-диагностики этой мутации в гете-ро- и гомозиготном состоянии, а также косвенной, по сцепленным с геном CFTR полиморфным генетическим маркерам любых других мутаций, являющихся причиной муковисцидоза.
Таким образом, на примере идентификации гена муковисцидоза удается показать все основные подходы, которые используют при идентификации гена наследственного заболевания, получившие название позиционного клонирования. По мере выполнения программы «Геном человека» необходимость использования такого подхода для идентификации му-тантного гена становится все меньше: после того как ген картирован, часто исследователям остается выбрать, какой из генов, уже картированных в этой области, можно рассматривать в качестве гена-кандидата и подтвердить предположение обнаружением мутаций в этом гене, которые, с одной стороны, не встречаются у здоровых людей, а с другой — могут быть функционально значимыми (этот подход называют картированием по положению).
В настоящее время картировано более 1,5 тыс. генов наследственных заболеваний (всего к концу февраля 2002 г. картировано почти 7800 генов человека). Более полные сведения можно получить из Online Mendelian Inheritance in Man (OMIM).
Картирование генов наследственных болезней, которое до недавнего времени было последним шагом в генетическом анализе, теперь стало отправной точкой на пути к идентификации гена. Далеко не всегда этот путь оказывается простым. Ген муковисцидоза, например, был идентифицирован только через 4 года после картирования, и таких примеров можно привести много. Однако в этом случае цель оправдывает время. Установление структуры гена позволяет предположить с большей или меньшей точностью, какой белок он кодирует, а следовательно, какова его физиологическая роль.
- Читать далее "Создание моделей наследственных болезней. Создание трансгенного животного."