Структура белка 4.1. Строение и кодирование белка 4.1
Этот глобулярный белок имеет Mw=78-80kD. Белок 4.1 состоит из 4 функционально различных доменов: основного N-концевого гидрофобного (Mw=30kD), гидрофильного (Mw=16kD), центрального (Mw=1 OkD) и карбокситерминального (Mw=22kD) [266,297,320]. N-концевой или мембрансвязывающий домен обеспечивает одно из прикрепляющих участков цитоскелета к липидному матриксу.
Благодаря наличию в его составе 36% ароматических остатков, а также аминокислот Не, Leu, Met и Val этот домен является высоко гидрофобным. Гидрофильный домен расположен между гидрофобным и спектринсвязывающим доменом и в своем составе содержит необычно высокий уровень Pro (9.3%).
Центральный или спектринсвязывающий домен является сильно заряженным благодаря содержанию в его составе до 46% Asp, Glu, Arg, Lys и His и форму длинной а-спирали. Карбокситерминальный домен является кислым и имеет короткую длину (117 аминокислот с общей Mw=12.636D).
При фракционировании в одномерном ПААГ-ЭФ белок 4.1 разделяется на 2 полосы: 4.1а (Mw=80kD) и 4.16 (Mw=78kD). По первичной структуре эти белки идентичны, а различную электрофоретическую подвижность можно объяснить дезаминированием белка 4.1а на Asn 502 и переходом его в низкомолекулярную форму на ранних посттрансляционных этапах биосинтеза. Низкомолекулярная форма доминирует в юных эритроцитах, а высокомолекулярная - в старых клеткак.
Белок 4.1 имеет связывающие участки не только для хвостовых частей спектриновых гетеродимеров, но и высокоаффинные сайты для глико-форинов и низкоаффинные - для АТБ, а также участки связывания для цитоскелетного белка р55 и белка pICIn. Белок 4.1 подвержен фосфорилированию на множестве сайтах различными типами протеинкиназ. Фосфорилирование на Туr в 627 позиции снижает способность белка 4.1 стабилизировать спектринактиновые комплексы.
Связывающая способность белка 4.1 с мембраной является тонко регулируемой. N-концевой домен имеет 2 цАМФ-зависимых участка фосфорилироваиия и 3 сайта для гликозилирования. Кроме гликозилирования, белок4.1 подвергается фосфорилированию протеинкиназой С, цАМФ-зависимой протеинкиназой, а также казеинкиназой I и II. На связывание белка 4.1 со спектринактиновой сетью существенное влияние оказывают полифосфоинозитиды, ДФГ, кальмодулин, концентрация катионов Са2+ и К+, содержание АТФ.
Снижение содержания белка 4.1 в мембране сопровождается уменьшением количества гликофорина С. Содержание белка 4.1 в мембране чувствительно к содержанию свободного Са2+ в цитоплазме эритроцита. При снижении внутриклеточной концентрации ионов Са2+ связывание белка 4.1 с мембранным скелетом снижено на 43%, тогда как увеличенная концентрация Са2+ в цитозоле сочетается со значительным дефицитом белка (83%).
- Вернуться в оглавление раздела "лабораторная диагностика"
Оглавление темы "Структура клеточных мембран":1. Клеточные мембраны. Плазматические мембраны
2. Виды биологических мембран. Разновидности клеточных липидов
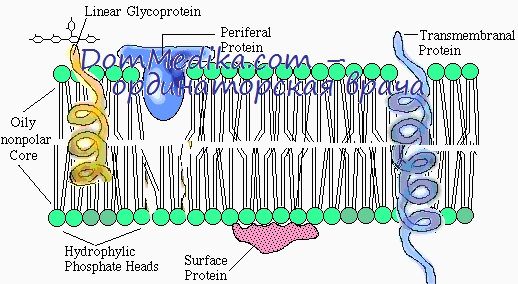
3. Мембранные белки. Структура мембранных белков клеток
4. Строение и функции белков клеточных мембран. Белки клеточных мембран эритроцитов
5. Периферические белки клеточных мембран. Спектрин клеточных мембран
6. Структура спектрина. Свойства спектрина клеточных мембран
7. Кодирование спектрина. Анкирин клеточных мембран
8. Содержание анкирина в клеточных мембранах. Анкирин в эритроцитах
9. Эритроцитарный анкириновый ген. Белок 4.1 клеточных мембран
10. Структура белка 4.1. Строение и кодирование белка 4.1