Антифагоцитарные факторы бактерий
Некоторые патогенные бактерии резистентны к действию бактерицидных факторов тканей хозяина благодаря функциям некоторых структурных элементов. Так, поли-D-глютаматная капсула Bacillus anthracis предохраняет бактериальную клетку от катионных белков (дефенсинов) сывороток и фагоцитов. Поверхностная мембрана грамотрицательных бактерий является проницаемым барьером, но не для гидрофобных веществ, таких как желчные кислоты, обычно токсичных для бактерий. Поверхностные липополисахариды (ЛПС) грамотрицательных бактерий предохраняют клетки от комплемент-опосредованного лизиса и действия лизоцима. Кроме того, большинство бактерий обладает добавочными факторами, которые помогают им выстоять в борьбе с клеточными и гуморальными факторами защиты организма хозяина.
Бактерии, которые захватываются и эффективно перевариваются фагоцитами, обычно не могут вызвать инфекционный процесс. Но патогенные бактерии в процессе эволюции выработали целый ряд механизмов, помогающих им не только избежать уничтожения фагоцитами, но и выживать и размножаться внутри них.
Как известно, процесс фагоцитоза включает следующие этапы: 1) контакт между фагоцитом и микробной клеткой; 2) поглощение; 3) образование фагосомы; 4) слияние фагосомы и лизосомы; 5) уничтожение и переваривание.
Чтобы избежать уничтожения в процессе фагоцитоза бактерии выработали специальные механизмы.
а) Подавление стадии поглощения. Некоторые бактерии продуцируют биологически активные вещества, способные препятствовать их поглощению фагоцитами. Как правило, это компоненты бактериальной клетки. Классическими примерами антифагоцитарных факторов являются: 1) полисахаридные капсулы Streptococcus pneumoniae, Neisseria meningitidis, Haemophilus influenzae, Klebsiella pneumoniae и Treponema pallidum; 2) М-белок и пили стрептококков группы А; 3) поверхностный полисахарид Pseudomonas aeruginosa; 4) О-антиген, ассоциированный с ЛПС энтеробактерий; 5) К-антиген (кислый полисахарид) Е. coli или Vi-антиген Salmonella typhi; 6) белок A Staphylococcus aureus, который связывается с Fc-доменом IgG и блокирует цитофильный домен антител. Таким образом, способность IgG реагировать как опсонин подавляется и опсонинопосредованное переваривание блокируется.
б) Выживание внутри фагоцита. Из внутриклеточных патогенов, к которым относятся Mycobacterium tuberculosis, Mycobacterium leprae, Listeria monocytogenes, представители родов Salmonella, Shigella, Yersinia, Brucella, Legionella, Rickettsia, Chlamydia, только некоторые, в том числе Mycobacterium tuberculosis, Mycobacterium leprae, Salmonella typhi и Listeria monocytogenes, способны выживать внутри макрофага или нейтрофила с вероятностью развития генерализованного инфекционного процесса (системной инфекции). У шигелл и дизентериеподобных энтеро-инвазивных кишечных палочек, проникающих и размножающихся в эпителиальных клетках, способность к пролиферации внутри профессиональных фагоцитов отсутствует. Следует отметить, что внутри фагоцитов бактерии защищены от активности гуморальных факторов защиты макроорганизма, в том числе комплемента, интерферонов и антител.
Большинство внутриклеточных патогенов имеет генетически детерминированные механизмы проникновения внутрь эпителиальных клеток (непрофессиональный фагоцитоз), обеспечивающие выживание внутри фаголизосом профессиональных фагоцитов. Так, клетки сальмонелл и иерсиний синтезируют различные белки наружной мембраны, часть из которых контролирует проникновение возбудителя внутрь эпителия, а другая — выживание внутри фагоцитов. Внутри фаголизосомы бактерии выживают посредством механизмов, которые блокируют бактерицидные вещества фагоцита. Так, клетки Legionella pneumophila включают в свою поверхность компонент комплемента С3b и используют этот белок для связывания бактерий с поверхностью макрофага. После поглощения бактерии остаются в вакуолях, которые не сливаются с лизосомами благодаря растворимому белку легионелл. То есть бактерии выживают внутри фагосом благодаря тому, что блокирована доставка лизосомального содержимого (смесь протеолитических ферментов) внутрь фагосом. Эта стратегия используется не только легионеллами, но и возбудителем туберкулеза, сальмонеллами и хламидиями. В случае микобактерий микроорганизмы синтезируют сульфатиды, которые выходят из фагосом и модифицируют лизосомальные мембраны таким образом, что слияние становится невозможно. Аналогичные вещества продуцируются легионеллами и хламидиями.
Некоторые бактерии обладают выраженной резистентностью к лизосомальному содержимому даже после слияния фагосомы и лизосомы. О факторах, которые обеспечивают такую резистентность, известно мало, однако примеры имеются. Mycobacterium leprae растет внутри фагосом даже после их слияния с лизосомами. Микобактерии имеют воскообразную гидрофобную клеточную стенку и миколевые кислоты (капсульный компонент), которые обеспечивают резистентность к лизосомальным ферментам. Похоже, что клеточные компоненты бруцелл интерферируют с бактерицидными механизмами фагоцитов.
Выживать внутри фагоцитов могут и внеклеточные патогены, например, S. pneumoniae, N. meningitides, N. gonorrhoeae, К. pneumoniae, Р. aeruginosa и др. Клетки стафилококков продуцируют каталазу и супероксиддисмутазу, способные нейтрализовать токсические кислородные радикалы, которые генерируются NADPH-оксидазой в фагоцитах. Компоненты клеточных мембран и капсул грамотрицательных бактерий (Salmonella, Yersinia, Brucella, патогенных Е. coli) могут предохранять пептидогликановый слой от литических ферментов. Бациллы Bacillus anthracis недоступны для переваривания за счет капсулы, построенной из поли-D-глютамата.
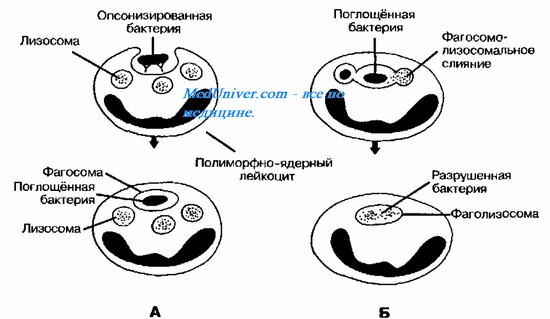
в) Выход из фагосом. Выход из фагосомальной вакуоли является решающим фактором для дальнейшего роста и размножения некоторых бактерий. Например, риккетсии входят в клетку в составе фагосом и уже через 30 с обнаруживаются в цитоплазме клетки. Это обеспечивается благодаря лизису фагосом ферментом фосфолипазой А. В случае листерий тот же эффект обеспечивается, по-видимому, посредством листериолизина О и двух форм фосфолипазы С. Оказавшись в цитоплазме, листерии двигаются по ходу полярной полимеризации актина. Шигеллы также лизируют фагосомы и индуцируют полимеризацию актина, что важно для внутриклеточного передвижения и межклеточного распространения возбудителя.
г) Вещества, убивающие или разрушающие макрофаги. Одной из ясных стратегий бактерий, такой, как защита против фагоцитоза, является прямая атака на профессиональные макрофаги. Вещества, способные вызывать гибель макрофагов, называют агрессинами. Большинство из них является ферментами или токсинами.
д) Разрушение макрофагов до поглощения. Многие грамположительные бактерии, особенно кокки, продуцируют внеклеточные вещества, которые убивают макрофаги. Некоторые из этих ферментов известны как гемолизины, названные так из-за их способности лизировать эритроциты. Например, патогенные стрептококки продуцируют гемолизин О, который связывается с холестерином на мембранах. Этот белок лизирует лизосомы, в результате чего их содержимое попадает в цитоплазму нейтрофила, что ведет к его гибели. Сходным механизмом обладает лейкоцидин стафилококков. Кроме гемолизинов, гибель макрофагов может наступить в результате действия экзотоксина А, продуцируемого синегнойной палочкой. Такие вещества, как отечный фактор возбудителя сибирской язвы и аденилатциклаза возбудителя коклюша резко снижают активность макрофагов.
е) Уничтожение фагоцитов после поглощения. Некоторые бактерии обладают токсическим действием на фагоциты после их поглощения. Они размножаются в фагосомах и выделяют вещества, которые проходят через мембрану фагосом и лизируют лизосомы. Такие внутриклеточные патогенны, как бруцеллы, листерии и сальмонеллы, в конечном итоге разрушают макрофаги, однако механизм этого разрушения не изучен.
ж) Природная видовая устойчивость макроорганизма. Некоторые виды животных имеют природную (генетическую) резистентность к определенным патогенным бактериям. Некоторые патогены инфицируют только человека, например, возбудители сифилиса, гонореи, краснухи, полиомиелита. В настоящее время имеется информация, позволяющая объяснить абсолютные видовые различия чувствительности к отдельным патогенам. К причинам этого феномена относятся:
1. Отсутствие специфических тканевых или клеточных рецепторов для прикрепления патогена. Например, способность различных штаммов энтеротоксигенных Е. coli колонизировать гастроинтестинальный тракт новорожденных, детей раннего возраста, телят, ягнят и поросят определяется наличием рецепторов для разных фимбриальных антигенов (разных факторов колонизации).
2. Температура тела и способность патогена расти при определенной температуре. Например, птицы не восприимчивы к Mycobacterium tuberculosis, так как эти бактерии не могут расти при высокой температуре. Вегетативные клетки Bacillus anthracis не могут расти в организме холоднокровных лягушек, но могут размножиться, если лягушка находится в помещении при 37 °С.
3. Отсутствие у хозяина определенных питательных веществ для роста бактерий. Так, пуринзависимые штаммы Salmonella enterica способны расти только в организмах, содержащих пурины. Мыши и крысы не имеют этих факторов, и пуринзависимые клоны сальмонелл в данном случае оказываются авирулентными. Однако введение пуринов в организм этих животных приводит к тому, что сальмонеллы начинают размножаться, т. е. становятся для них вирулентными.
4. Отсутствие мишеней для токсинов. Большинство токсинов проявляет свой токсический эффект только после связывания с чувствительными клетками организма. Однако некоторые животные не имеют таких рецепторов и поэтому приобретают устойчивость к токсическому действию. Например, дифтерийный токсин не способен убивать крыс и удаляется из организма с мочой, а последняя при этом приобретает свойства, токсичные для морских свинок.
- Читать далее "Грызуны используемые в качестве лабораторных животных в микробиологических лабораториях"
Редактор: Искандер Милевски. Дата публикации: 9.08.2019