Молекулярная организация (строение) бактериальных токсинов
Обычно бактериальные токсины белковой природы имеют субъединичную структуру (A-В). Это означает, что молекула токсина состоит из двух компонентов, которые отличаются друг от друга по структурным, функциональным и иммунохимическим свойствам. Указанные компоненты для краткости называют А и В, причем компонент А обычно выполняет энзиматическую функцию, а компонент В — рецепторсвязывающую. В дальнейшем компонет А будет обозначаться как Р-компонент (Р-регуляторный), а компонент В — как РС-компонент (РС-рецепторсвязывающий).
В общем плане эти токсины представляют собой химеры, состоящие из фермента и лектина, причем каждый из них в отдельности не обладает токсической активностью. В то же время токсическая функция является гиперфункцией динамически подвижной цельной молекулы. В результате этой гиперфункции происходит перенос энзиматического участка молекулы внутрь клетки и затем — нарушение ее жизнедеятельности. Понятно, что наличие как минимум двух различных белков в молекуле токсина является отражением молекулярной организации генов, кодирующих эту молекулу.
а) Рецепторсвязывающие компоненты бактериальных токсинов. Регуляторный компонент функционирует не в клетке, которая его синтезирует, а в гетерологичных клеточных системах. Это достигается благодаря следующим функциям рецепторсвязывающих компонентов:
1. Функция белок-углеводного или белок-белкового узнавания, т.е. распознавание и взаимодействие с рецепторами чувствительных клеток. Здесь следует отметить, что со стороны клеток в качестве рецепторов выступают структуры, которые используются не только токсинами, но и гормонами и интерферонами.
2. Функция транслокации регуляторного компонента во внутренние компартменты гетерологичных или чувствительных клеток. В пользу этого говорят данные о том, что P-компоненты являются нетоксичными или биологически инертными белками при взаимодействии их с чувствительными клетками в отсутствии РС-компонентов.
3. Функция транспорта токсических молекул из клеток, в которых они синтезируются.
4. И, наконец, экранирующая функция. В составе токсической молекулы рецепторсвязывающие компоненты блокируют активность энзиматического компонента. Для активации молекулы необходим протеолиз и диссоциация компонентов.
Таким образом, рецепторсвязывающие компоненты бактериальных токсинов выполняют ряд вспомогательных функций, наиболее важной из которых является доставка регуляторного компонента к клеткам-мишеням, а в случае наличия в их составе транслокационного домена—перенос регуляторного компонента во внутреннюю среду чувствительных клеток.
б) Свойства регуляторных компонентов бактериальных токсинов. Известно четыре основных типа реакций, характерных для токсических молекул: АДФ-рибозилирование, осуществляемое АДФ-рибозилтрансферазой целого ряда токсинов; гидролиз рРНК, который используют шигеллоподобные токсины 1 и 2 типов; генерация вторичных мессенджеров, осуществляемая отечным фактором токсина сибиреязвенного микроба, а, возможно, и Bacillus cereus; индукция протеинкиназы ST-подобными токсинами кишечных бактерий.
Ковалентная модификация внутриклеточных белков путем АДФ-рибозилирования является одним из ключевых механизмов регуляции клеточных процессов в природе. Имеющиеся в литературе данные свидетельствуют, что во многих случаях компоненты А бактериальных токсинов являются АДФ-рибозилтрансферазами. Эти токсины вырабатываются различными видами бактерий, которые либо находятся в близком родстве, как, например, сальмонеллы, энтеропатогенные кишечные палочки, холерные вибрионы и т. д., либо весьма далеки друг от друга, например, в случае клостридий или коринебактерий. Однако для всех этих токсинов молекулярной реакцией поражения внутриклеточных мишеней является реакция АДФ-рибозилирования.
В результате рибозилирования внутриклеточный белок теряет способность участвовать в том или ином жизненно важном процессе клетки, что ведет либо к гибели последней, либо к нарушению ее нормального функционирования.
Если верно мнение, что бактериальные токсины используют реакции, связанные с нормальной регуляцией клетки, то в клетке должен иметься регулятор, который осуществляет эту функцию. По крайней мере в некоторых случаях это уже показано. Например, P-компоненты холерного энтеротоксина, LT и LT-подобного токсинов кишечной палочки представляют собой аргининспецифические АДФ-рибозилтрансферазы, которые рибозилируют регуляторную субъединицу аденилатциклазы, что приводит к устойчивой ее активации.
Фермент с аналогичной функцией обнаружен в эритроцитах человека и других клетках млекопитающих; было показано, что он также рибозилирует Ni-белок, что приводит к активации аденилатциклазы клеток мозга.
Известен и другой пример. Так, P-компоненты дифтерийного токсина и экзотоксина A Pseudomonas aeruginosa являются дифтамидспецифическими АДФ-рибозилтрансферазами, которые рибозилируют (переносят аденозиндифосфатрибозу от НАД на белок-мишень) фактор элонгации 2 и, таким образом инактивируя его, подавляют биосинтез белка в клетках. Аналогичный фермент обнаружен в целом ряде эукаритических клеток. При этом показано, что он так же, как дифтерийный и синегнойный токсины, рибозилирует фактор элонгации 2.
Обнаруживается поразительное сходство между трансферазами про- и эукариотического происхождения, которые активируют аденилатциклазу (фермент, генерирующий вторичный мессенджер циклический аденозин мононуклеотид). Прежде всего это касается молекулярной массы, констант Михаэлиса для различных субстратов, количества оборотов, стереохимичности субстратного продукта, акцептора АДФ-рибозы, внутриклеточной мишени и конечного биологического эффекта на клетку.
Другим известным механизмом действия токсинов на молекулярном уровне является нарушение биосинтеза белка посредством инактивации рибосом. Так действуют токсин шига и шигаподобные токсины 1 и 2 типов, которые представляют собой N-гликозидазы. P-компоненты этих токсинов энзиматически расщепляют N-гликозидную связь у аденина в положении 4324 28S рибосомальной ДНК. В результате происходит отщепление 400 нуклеотидов с 3’-конца рРНК, что служит препятствием для присоединения аминоацил-tRNA.
Следующий механизм — генерация вторичных мессенжеров. В этом случае в токсическую молекулу включен фермент аденилатциклаза, который генерирует вторичный мессенжер — цАМФ. Аденилатциклаза, или отечный фактор Bacillus anthracis, не только по кинетическим, но и по антигенным свойствам имеет сходство с кальмодулинзависимой аденилатциклазой эукариотических клеток.
Таким образом, в основе токсического действия бактериальных токсинов лежит нарушение регуляции важных биохимических процессов клетки, и нарушение этих процессов происходит за счет функции P-компонентов токсических молекул, которые по своей природе являются регуляторными элементами.
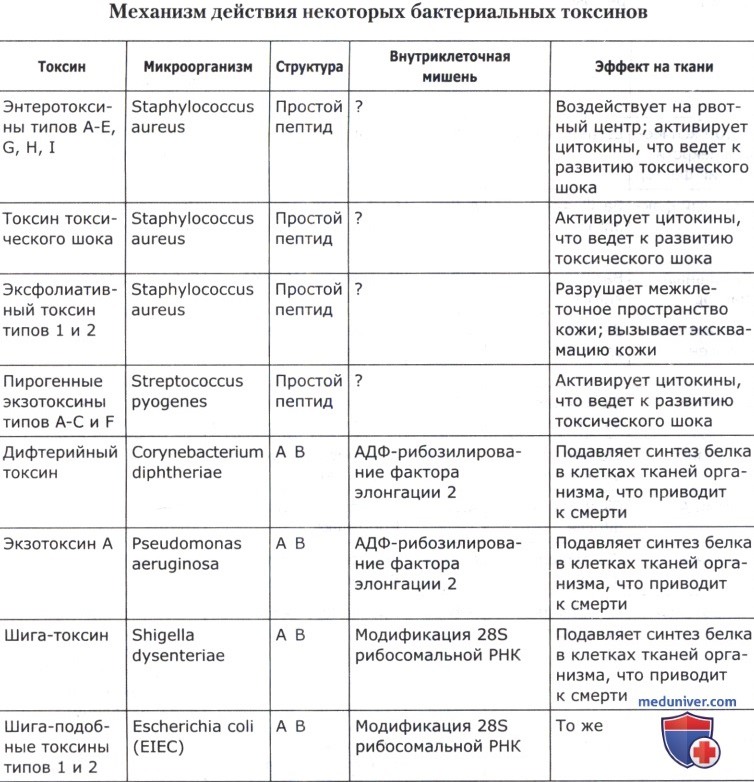
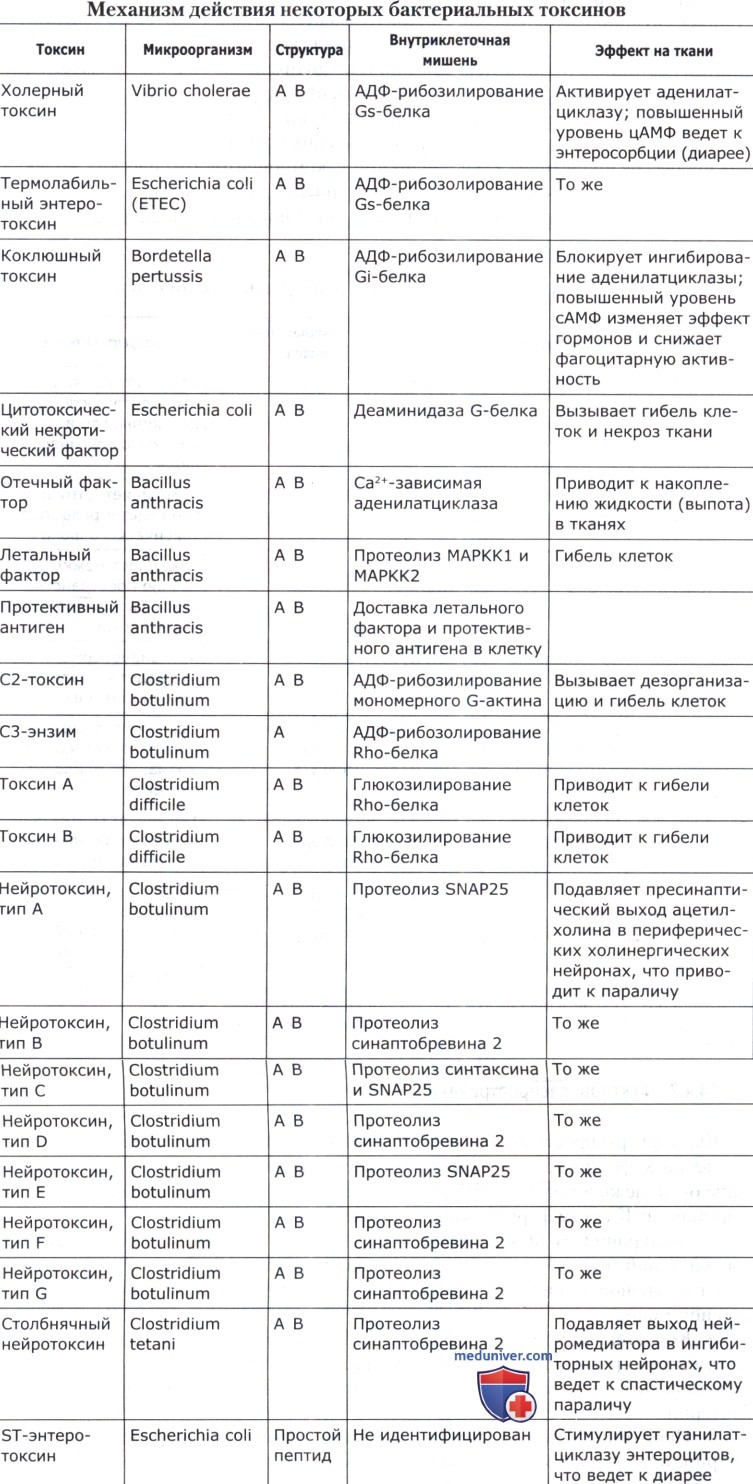
Механизм действия ряда бактериальных токсинов приведен в таблице ниже.
- Читать далее "Сидерофоры бактерий"
Редактор: Искандер Милевски. Дата публикации: 5.08.2019
- Патогенность и вирулентность микроорганизмов
- Факторы патогенности бактерий
- Формирование патогенных штаммов микроорганизмов
- Геномные острова патогенности
- Факторы адгезии и колонизации бактерий
- Факторы инвазивности бактерий
- Факторы патогенности бактерий с токсической функцией
- Молекулярная организация (строение) бактериальных токсинов
- Сидерофоры бактерий