В-лимфоциты лимфатических узлов - созревание, функции
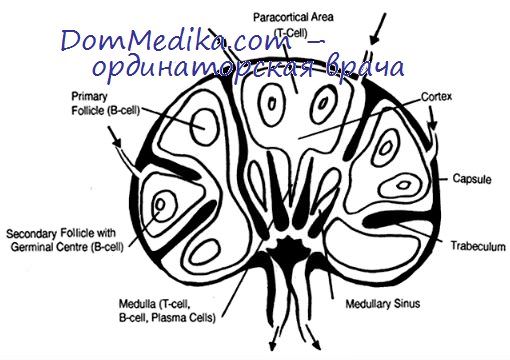
В лимфатических узлах В-лимфоциты сосредоточены в основном в фолликулах. Первичные фолликулы и зона мантии вторичных фолликулов состоят из малых лимфоцитов, большинство из которых характеризуется отсутствием признаков активации. Чаще всего эти клетки имеют изотип IgM+IgD или IgM; антигены CD23, CD5, CD10, CD38, как правило, отсутствуют. Лимфоциты экспрессируют на мембране CD 19, CD20, HLA-DR, CD2I, CD22, CD24, CD37 и ряд других маркеров зрелых В-лимфоцитов, т. е. их иммунологический фенотип является характерным для периферических В-клеток.
У плода человека и новорожденных фенотип лимфоцитов первичных фолликулов лимфатических узлов и селезенки характеризуется экспрессией молекулы CD5, что отличает эти клетки от клеток первичных фолликулов тех же органов взрослых. Содержание клеток CD5+ в лимфатических узлах у взрослых невелико и составляет 2— 3 % от всех В-лимфоцитов.
Наивные В-лимфоциты поступают в лимфатический узел и другие периферические лимфоидные органы с током крови через высокий эндотелий венул, расположенных в паракортикальной зоне. Первичная активация В-клеток происходит именно в этих Т-клеточных областях периферических лимфоидных органов: паракортикальной зоне лимфатических узлов и лимфоидной ткани слизистых оболочек, периартериолярных лимфоидных муфтах селезенки. Последствия связывания иммуноглобулино-вых рецепторов В-лимфоцитов с антигеном во многом зависят от свойств самого антигена.
Некоторые антигены (так называемые тимуснезависимые) способны вызывать пролиферацию и дифференцировку В-клеток без помощи Т-лимфоцитов. Тимуснезависимые антигены I типа являются поликлональными активаторами, а тимуснезависимые антигены II типа — это, как правило, полисахариды со множеством регулярно повторяющихся одинаковых антигенных детерминант, способных перекрестно связывать мембранные IgM В-лимфоцитов и вызывать активацию этих клеток.
Активация В-клеток под действием тимусзависи-мых антигенов (чаще это белки, нуждающиеся в переработке — процессинге и комплексировании с молекулами HLA для эффективного распознавания Т-лимфоцитами) происходит при участии Т-хелперов и дендритических клеток в паракортикальной зоне. В-лимфоциты взаимодействуют с CD4+ Т-хелперными клетками, активированными антигенными производными, представленными в комплексе с молекулами HLA II класса на интердигитирующих клетках.
При активации хелперных клеток молекула CD28 на СВ4+-лимфоцитах связывается с ее лиган-дом CD80 на интердигитирующих клетках. Взаимодействие Т- и В-лимфоцитов осуществляется двумя способами: контактным (клетка-клетка) и с помощью цитокинов. В контактных взаимодействиях принимают участие со стороны В-клеток молекулы CD40, LFA-1, LFA-3 и комплементарные им структуры Т-лимфоцитов — лиганд CD40 (появляется на активированных Т-клетках), ICAM-1 и CD2. Основными цитокинами, синтезируемыми Т-хелперными лимфоцитами и поддерживающими активацию и пролиферацию антигенспецифичных В-клеток, являются ИЛ-4, а также ИЛ-5 и ИФН-у.
Отражением процесса активации служат появление на мембране В-лимфоцита молекулы CD23, повышенная экспрессия молекул HLA II класса, а затем утрата мембранного IgD. Особая группа антигенов, включающая главным образом аутологичные антигены и немногочисленные (преимущественно тимуснезависимые) экзогенные антигены, взаимодействуя с В-лимфоцитами, ведет к появлению экспрессии CD5 в процессе активации. В-лимфоциты CD5-линии используют ограниченный репертуар Vh-генов.
Появление CD5-антигена происходит лишь на 20 % В-лимфоцитов и достигает пика через 60 ч после активации, при этом клетки еще не пролиферируют, а остаются в фазе G0—G1 клеточного цикла. Именно CD5-позитивные В-лимфоциты экспрессируют антиген зрелых В-клеток CD39. В процессе активации В-лимфоцитов, сопровождающейся экспрессией CD5, на мембране клеток появляется молекула CD38 (экспрессия этих молекул увеличивается и при активации CD5-позитивных В-лимфоцитов). ИЛ-4 предотвращает появление на мембране В-лимфоцитов CD5 (и ассоциированное с этим появление CD38) в процессе активации.
При активации наивных В-клеток пуповинной крови появление CD5 также происходит на 2—3-й день, а к 5-му дню антиген полностью исчезает. Однако в этом случае экспрессия CD5 не сопровождается увеличением экспрессии CD38 на мембране клеток.
Эти данные убедительно свидетельствуют о том, что определенная субпопуляция В-лимфоцитов при активации способна к экспрессии CD5 на мембране. Именно эти клетки являются опухолевым субстратом лимфомы из малых лимфоцитов/хронического лимфолейкоза. Изменения иммунофенотипа этих клеток в динамике в процессе активации свидетельствуют о том, что при появлении на мембране антигенов, свойственных клеткам зародышевых центров, молекула CD5 утрачивается. Подобные наблюдения хорошо коррелируют с иммуноморфологическими данными, свидетельствующими об отсутствии экспрессии CD5 на клетках зародышевых центров.
В ходе иммунного ответа на тимусзависимые и тимуснезависимые антигены активированные В-лимфоциты могут далее дифференцироваться в плазматические клетки, синтезирующие антитела IgM, или дать начало реакциям зародышевых центров.
Следует обратить внимание на особенности терминологии. Понятие «первичный иммунный ответ» применимо обычно к ответу Т- или В-лимфоцитов на первый контакт с антигеном, т. е. наблюдается в неиммунном к данному антигену организме. Термин «первичный иммунный ответ» как бы противопоставляется «вторичному иммунному ответу», т. е. более быстрому ответу В-клеток памяти на повторный контакт с антигеном. Первичный иммунный ответ — это ответ наивных зрелых В-клеток (IgM+, lgD+) на первый контакт с антигеном. Связывание мембранными иммуноглобулинами, входящими в состав BCR В-лимфоцитов, специфического антигена ведет к их переходу в бластную форму и пролиферации.
Эти экстрафолликулярные (процесс происходит в паракортикальной зоне) крупные В-бласты дают потомство в виде двух типов клеток — короткоживущих плазматических клеток и В-клеток, приобретающих способность давать начало зародышевым центрам. Короткоживущие плазматические клетки обеспечивают продукцию IgM в ходе первичного иммунного ответа. Другие потомки экстрафолликулярных В-бластов поступают в фолликул и проходят дифференцировку в зародышевом центре.
Вторичный гуморальный иммунный ответ осуществляется, как правило, антителами IgG, IgA или IgE изотипов, а первичный — преимущественно антителами класса IgM. Вместе с тем в ходе первичного иммунного ответа на сложные антигены (например, эритроциты барана, как это показано в экспериментах на мышах) также существует несколько фаз. Фаза 1: активация и деление лимфоцитов через 1— 2 дня после иммунизации. Частота митозов Т-лимфоцитов становится максимальной примерно на 3-й день, а для В-лимфоцитов — примерно на 4-й день.
Фаза 2. антителообразующие клетки, преимущественно IgM, появляются в лимфатических узлах на 3—4-й день и вскоре становятся основным компонентом мякотных тяжей. Фаза 3: на 4—5-й день (после появления сывороточных антител) обнаруживаются зародышевые центры. Первичный IgM-антительный гуморальный иммунный ответ протекает без формирования зародышевых центров фолликулов лимфатических узлов. Фаза 4: на 5—7-й день происходит нарастание сывороточных титров IgG. Фаза 5: на 9—15-й день нарастают титры сывороточных IgA. Таким образом, формирование зародышевых центров с переключением классов иммуноглобулинов и образованием клеток памяти — это 2-я фаза (первая — продукция IgM без формирования зародышевых центров) в ходе реализации иммунного ответа на первичный контакт с антигеном.
- Читать "Внутрифолликулярная дифференцировка В-лимфоцитов"
Оглавление темы "Клетки крови":- Онтогенез Т-лимфоцитов - образование
- Созревание (онтогенез) NK-клеток
- Созревание (онтогенез) лимфоидных дендритных клеток Лангерганса
- Рециркуляция лимфоцитов в организме
- Лимфатические узлы - формирование Т-лимфоцитов
- Функции Т-лимфоцитов лимфатических узлов
- В-лимфоциты лимфатических узлов - созревание, функции
- Внутрифолликулярная дифференцировка В-лимфоцитов
- Селезенка - строение, формирование Т- и В-лимфоцитов
- Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге