Монокарбоновые субстраты. Перенос катионов в матрикс митохондрий
Большинство монокарбоновых субстратов (например, ацетат), по-видимому, обладает способностью свободно диффундировать сквозь внутреннюю мембрану без участия специфических систем переноса. Однако для облегчения перемещения высших жирных кислот из цитоплазмы, где они образуются в ходе реакций липолиза, в матрикс, где находятся ферменты окисления жирных кислот, существует специальный механизм переноса. Перенос алифатических ацильных групп сквозь митохондриальную внутреннюю мембрану сопровождается образованием диффундирующего эфира с соединением, называемым карнитином. Эту реакцию катализирует специальный фермент, встроенный во внутреннюю мембрану митохондрий и катализирующий процессы, протекающие в определенных направлениях (изнутри наружу и наоборот). Процесс, направленный наружу, сводится к переносу ацильной группы (остатка жирной кислоты), находящейся в виде активированного эфира КоА, на карнитин, располагающийся на наружной, обращенной к цитоплазме поверхности внутренней мембраны:
Ацил-КоА + Карнитин = Ацилкарнитин + КоА.
Процесс, направленный внутрь, заключается в ферментативном обращении предыдущего процесса, т. е. в превращении ацилкарни-тина, обладающего свойством свободно проходить сквозь мембрану, в ацил-КоА на той стороне внутренней мембраны митохондрий, которая обращена в сторону матрикса.
Важно отметить, что этот механизм переноса обеспечивает эффективный транспорт ацил-КоА, который не способен проникать сквозь внутреннюю мембрану митохондрий, из цитоплазмы в матрикс без изменения содержания кофермента А в этих двух отсеках клетки.
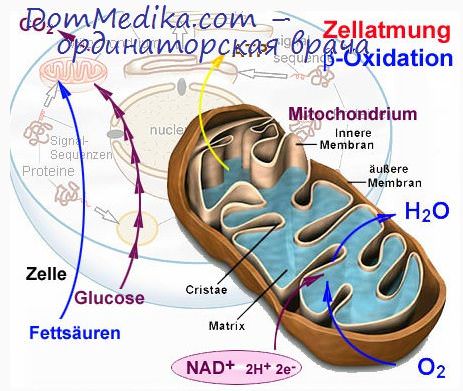
Перенос катионов в матрикс митохондрий непосредственно связан с передачей энергии внутренней мембраны митохондрий На основе представлений о возникновении электродвижущих сил, сопровождающих движение ионов сквозь эту мембрану, вероятно, можно будет понять в конечном счете механизм запасания энергии при окислительном фосфорилировании. Прежде всего следует отметить, что внутренняя мембрана митохондрий совершенно непроницаема для большинства катионов вообще и для ионов водорода в частности. Поэтому и возможно возникновение ионного градиента между содержимым митохондрий и цитоплазмой.
Если возникновение такого градиента обусловлено перемещением Н+ в определенном направлении, то создается протон-движущая сила, или трансмембранный градиент рН. Более того, совершенно ясно, что переносчики электронов дыхательной цепи ориентированы во внутренней мембране митохондрий асимметрично, а именно так, что они доступны для соответствующих субстратов либо с одной стороны мембраны, либо с другой ее стороны. Например, НАД-Н может взаимодействовать с дыхательной цепью только со стороны матрикса, или с М-стороны, тогда как цитохром с функционирует с наружной, цитоплазматической поверхности, или с Ц-стороны. Такое расположение переносчиков электронов во внутренней мембране определяет движение электронов при окислительно-восстановительных ре акциях дыхательной цепи в строго определенном направлении.
Следовательно, процесс переноса электронов может привести к разделению зарядов и возникновению ионных градиентов между митохондриями и цитоплазмой, подобно тому как контролируемое сгорание водорода в топке приводит к выработке электрической энергии в результате разделения заряженных Н+- и ОН-ионов, возникающих при окислительной реакции: 2Н2 + 02=>2Н++20Н.
При тканевом дыхании протоны выталкиваются из митохондрий, оставляя обращенную к матриксу поверхность внутренней мембраны заряженной отрицательно, тогда как сам матрикс оказывается щелочным по сравнению с окружающей средой, т. е. цитоплазмой Этот выброс Н+ свидетельствует о такой направленности расположения переносчиков электронов, которая обеспечивает освобожжение Н+ в окружающую среду, тогда как захват Н+ происходит при цитохромной реакции на обращенной к матриксу поверхности мембраны:
2Fea+-цитохром а3 + 2Н+ +1/202 — 2 Ре3+-цитохром а3 + Н20.
Таким образом, возникающий в результате окислительных реакций в дыхательной цепи повышенный энергетический уровень митохондрий можно выразить в терминах электрохимического градиента Н4; в сущности, присущая данному субстрату энергия восстановительного потенциала превращается в электрохимический потенциал.
- Читать "Обмен кальция в митохондриях. Компоненты механизма сопряжения митохондрий"
Оглавление темы "Энергообмен в тканях":1. Митохондрии. Значение митохондрий в обмене веществ
2. Циркуляция субстратов. Транспортеры
3. Монокарбоновые субстраты. Перенос катионов в матрикс митохондрий
4. Обмен кальция в митохондриях. Компоненты механизма сопряжения митохондрий
5. Феномен разобщения. Энергозависимые системы переноса
6. Функции всасывания и секреции. Активный перенос веществ
7. Энергетический обмен в нервной ткани. Возбудимость клеток
8. Распространение потенциала действия по нерву. Креатинфосфат
9. Энергообмен в головном мозгу. Энергетический обмен в мышечной ткани
10. Миоглобин. Энергообмен в красных и белых мышцах