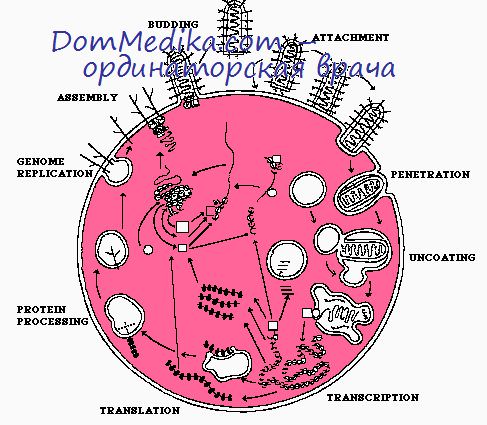
Метаболизм пораженных рабдовирусом клеток. T-частицы рабдовирусов
Степень подавления различными рабдовирусами клеточных синтезов сильно варьирует. Она минимальна, например, при персистентных инфекциях, вызванных вирусом бешенства, когда зараженные клетки продолжают производить вирус практически неопределенно долго. Вместе с тем ВВС, который и с этой точки зрения является единственным тщательно изученным рабдовирусом, способен подавлять синтез клеточных макромолекул.
При высокой множественности заражения подавление синтеза РНК и белка клетки-хозяина вызывают как инфекционные, так и инактивированные ультрафиолетом В- или Т-частицы ВВС (Хуанг и Вагнер, 1965; Вагнер и др., 1970). Последнее, возможно, объясняется токсическим действием белков инокулума, поскольку известно, что РНК, входящая в состав УФ-облученных частиц, не функционирует. Существует и другой механизм подавления клеточного белкового синтеза, который, вероятно, зависит от образования вирусного белка (Верц и Янгнер, 1972). Это подавление вызывается лишь живым вирусом и не происходит в присутствии циклогексимида. Довольно неожиданны данные о том, что мелкобляшечный мутант подавляет клеточный белковый синтез быстрее (90% за 2 ч), чем крупнобляшечный (90% за 8 ч). Хуанг и др. (1970), показали, что на третьем часу после заражения использованным штаммом ВВС происходит разрушение полисом, после чего образуется новая популяция более мелких вирус-специфических полисом, количество которых достигает максимума на четвертом часу после начала инфекции.
T-частицы рабдовирусов
Т-частицы (truncated — усеченный) ВВС, которые описали Купер и Белет (1969), а затем Хэкетт (1964), Стемпфер и др. (1971), обозначают как ДИ (дефектные интерферирующие)-Т-частицы. При таком обозначении подчеркиваются два характерных свойства этих структур: абсолютная неинфекционность в отсутствие инфекционных В-частиц и способность специфически препятствовать репликации В-частиц. ДИ-Т-частицы, описанные в литературе, в основном имеют длину, равную '/з длины В-частиц, однако по современным представлениям размеры частиц могут широко варьировать в зависимости от штамма вируса и типа клеток (обзор Хуанг и Балтимора, 1970).
Превек и Канг (1970) описали два класса Т-частиц, образуемых при использовании полученного ими штамма ВВС, причем успешно интерферировать с гетерологичным штаммом могли лишь частицы, принадлежащие к классу более длинных частиц. Длина Т-частицы — характерное свойство родительского штамма в случае fc-мутантов Прингла (Райхман и др., 1971). Пассирование без разведения некоторых из этих мутантов при разрешающей температуре приводит к преимущественному образованию ДИ-Т-частиц. Существуют определенные типы клеток, в которых аутоинтерференция ДИ-Т-частиц проявляется легче, чем в других (Перро и Холленд, 1972).
Клонированные В-частицы не образуют Т-частиц примерно до третьего пассажа с высокой множественностью заражения (Стемпфер и др., 1971). При последующих пассажах относительное количество Т-частиц возрастает, а урожай В-частиц в результате интерференции пропорционально падает. Суммарный синтез вирусного белка (Мадд и Саммерс, 1970а; Вагнер и др., 1970) и вирусных РНК (Стемпфер и др., 1969) подавляется. Однако транскрипция РНК В-частиц не изменяется, если до заражения В-частицами клетки обрабатывают Т-частицами в присутствии циклогексимида (Хуанг и Мэндерс, 1972).
Для интерференции необходима РНК ДИ-Т-частиц; инактивированные ультрафиолетом частицы интерферировать не способны (Хуанг и Вагнер, 1966а). Несмотря на это, Т-частицы не обладают транскриптазной активностью in vitro (Бишоп и Рой, 1971) и не синтезируют РНК in vivo (Стемпфер и др., 1969). Очевидно, транскрипция РНК Т-частиц не является обязательным условием для осуществления интерференции. Это заключение находит косвенное подтверждение в следующем наблюдении: хотя ДИ-Т-частицы любого ts-мутанта Прингла мешают размножению В-частиц дикого типа, ни один из этих мутантов не способен комплементировать с В-частицами /s-мутантов из других групп комплементации (Райхман и др., 1971). Более того, Сривэлсэну (1970b) удалось вызвать интерференцию, используя очищенную РНК из Т-частиц. Таким образом, принятая в настоящее время гипотеза заключается в том, что РНК Т-частиц, эффективно связываясь с репликазой ВВС, специфически интерферирует с репликацией (но не с транскрипцией) РНК В-частиц (Стемпфер и др., 1969; Сривэлсэн, 1970).
- Вернуться в оглавление раздела "Вирусология"
Оглавление темы "Характеристика парамиксовирусов и рабдовирусов":1. Метаболизм клетки пораженной вирусом гриппа. Зависимость вируса гриппа от клеток хозяина
2. Неполный вирус гриппа. Парамиксовирусы
3. Ранняя стадия парамиксовирусной инфекции. Транскрипция парамиксовирусов
4. Трансляция и репликация РНК парамиксовирусов. Сборка и выход парамиксовирусов из клетки
5. Метаболизм пораженных парамиксовирусами клеток. Слияние клеток пораженных парамиксовирусами
6. Размножение коронавирусов человека. Размножение аренавирусов и супергруппы Буниамвера
7. Вирус везикулярного стоматита. Цикл размножения и транскрипция рабдовирусов
8. Трансляция рабдовирусов. Репликация РНК рабдовирусов
9. Сборка и выход рабдовирусов из клетки. Степень выраженности генов рабдовирусов
10. Метаболизм пораженных рабдовирусом клеток. T-частицы рабдовирусов