Влияние интерферона на трансляцию. Механизм воздействия интерфенонов на вирусы
На вирусные частицы интерферон непосредственно не действует; не влияет он и на их адсорбцию или проникновение в клетку. Однако неясно, действует ли интерферон прямо на клеточную мембрану или же он должен сначала проникнуть в клетку. При обработке клеток интерфероном концентрация его в среде почти не уменьшается (Баклер и Бэрон, 1966).
Эффективность интерферона в отношении индукции резистентного состояния зависит от его концентрации, а не от абсолютного количества (Бэрон и др., 1967); это позволяет предполагать, что перенос интерферона внутрь клетки зависит от его концентрации. Интерферон может связываться клетками на холоду; обработка таких клеток трипсином перед их согреванием до 37 °С предотвращает развитие резистентности (Фридмен, 1967). Уже довольно давно высказывались предположения, что интерферон индуцирует синтез нового белка, который подавляет синтез вирусных макромолекул, не влияя на нормальную синтетическую активность клетки (Тейлор, 1964, 1965; Локхарт, 1964). Такое подавление могло бы происходить на уровне образования или трансляции мРНК.
Несмотря на значительные усилия, затраченные на изучение этого вопроса, точный механизм действия интерферона еще однозначно не установлен. Недавно было опубликовано несколько обширных обзоров (Леви и др., 1970; Колби и Морган, 1971), и мы, не пытаясь охватить проблему полностью, ограничимся рассмотрением отдельных работ, в которых представлены данные о механизме действия интерферона.
Некоторые указания на то, что интерферон действует на уровне трансляции, были получены Маркусом и Сэлбом (1966), которые сравнили взаимодействие меченой вирусной и клеточной РНК с рибосомами из фибробластов куриного эмбриона, обработанных и не обработанных интерфероном. В опытах без интерферона РНК вируса Синдбис связывалась при низкой температуре с рибосомами этих фибробластов. После повышения температуры эти РНК-рибосомные комплексы распадались и одновременно происходило включение аминокислот в полипептиды.
Но если источником рибосом были клетки, обработанные интерфероном, РНК-рибосомные комплексы по-прежнему формировались на холоду, но не подвергались распаду при нагревании. Авторы пришли к выводу, что рибосомы из клеток, обработанных интерфероном, могут связываться с вирусной РНК, но не транслируют ее, хотя их способность транслировать мРНК, выделенную из полисом клетки-хозяина, сохраняется.
Мягкая обработка таких рибосом трипсином восстанавливала их способность транслировать вирусную РНК (Маркус и Сэлб, 1968), и авторы предположили, что интерферон индуцирует синтез закодированного в клеточном геноме «белка, подавляющего трансляцию», который связывается с рибосомами и ингибирует трансляцию ими вирусной РНК.
Керр и его сотрудники (Керр и др., 1970; Керр, 1971) провели тщательную проверку этих результатов на такой же системе, с какой работали Маркус и Сэлб. Они искали такие изменения в белках рибосомной фракции, которые могли бы указывать на присутствие гипотетического ингибитора. Эти поиски дали отрицательный результат; авторы не смогли также воспроизвести наблюдавшееся Маркусом и Сэлбом подавление трансляции.
Керр и сотр. (1970) нашли, что распад РНК-рибосомных комплексов, описанный Маркусом и Сэлбом, не имеет отношения к включению аминокислот и поэтому не может быть надежным показателем трансляции. В отличие от результатов Маркуса и Сэлба рибосомы из обработанных интерфероном и из контрольных клеток ничем не различались по способности транслировать полиуридиловую кислоту и подвергаться распаду во время включения аминокислот.
Тем не менее работа Маркуса и Сэлба принесла пользу тем, что в ней была четко сформулирована рабочая гипотеза относительно механизма действия интерферона, побудившая исследователей к ее экспериментальной проверке. Эта модель остается приемлемой в принципе, но не в деталях, касающихся действия антивирусного белка. Следует также иметь в виду, что еще нет прямых данных о существовании антивирусного белка, образующегося под действием интерферона.
Если такой белок существует, то ген, ответственный за его синтез, локализован не в той хромосоме, в которой находится генинтерферона; это установили Кассинджена и сотр. (1971), изучавшие образование и действие интерферона в гибридных клеточных линиях.
Результаты Керра не опровергают гипотезу о модификации рибосом; они указывают на то, что изменение в обработанных интерфероном клетках, возможно, связано с каким-то фактором, необходимым для синтеза вирусного белка, — фактором, который сам не является частью рибосомы, но способен соединяться с нею при определенных условиях.
Позднее Керр (1971) сообщил о частичном подавлении трансляции РНК вируса энцефаломиокардита рибосомами из клеток, обработанных интерфероном. Этот результат воспроизводился только тогда, когда источником рибосом служила определенная фракция клеточных микросом. Из полученных данных не видно, на каком уровне происходит ограничение трансляции — на уровне инициации, образования пептидных связей или освобождения полипептида. Для дальнейшего прогресса в этой области необходимо определить число и природу факторов, контролирующих трансляцию вирусной РНК.
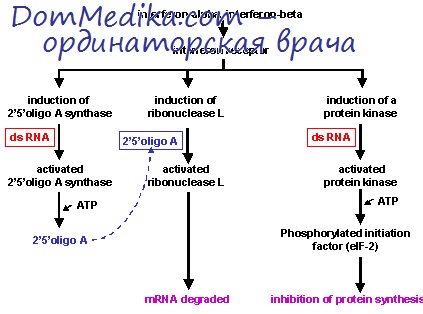
- Читать далее "Точки приложения интерферона в вирусах. Как ИФН тормозит вирусы?"
Оглавление темы "Эффекты интерферонов. Воздействие вирусов на клетки":1. Влияние интерферона на трансляцию. Механиз воздействия интерфенонов на вирусы
2. Точки приложения интерферона в вирусах. Как ИФН тормозит вирусы?
3. Влияние интерферона на транскрипцию вирусов. Торможении транскрипции интерферонами
4. Усиление вирусов интерферонами. Репродукция при воздействии интерферонов
5. Усиление размножения РНК вирусов. Воздействие вируса на макроорганизм
6. Механизмы повреждения клеток вирусами. Гибель клеток от вирусов
7. Подавление синтеза в клетках вируса. Цитопатические эффекты вирусов
8. Гистологические изменения зараженных клеток. Тельца включения в зараженных клетках
9. Слияние зараженных вирусами клеток. Механизмы слияния клеток
10. Вирусные гемолизины. Митоз при вирусных инфекциях