Мембранный потенциал: потенциал покоя, потенциал действия, нервный импульс, рефрактерный период
Биологические мембраны в большинстве своем являются полупроницаемыми, поскольку в них встроены ионные каналы. Мы будем называть их просто мембранами. Электрическая активность нейрона обусловлена движением специфических ионов через мембрану нейрона по специальным каналам. Именно это позволяет волнам электрической активности распространяться по мембранам, обеспечивая передачу информации в нервной системе. От чего же зависит транспорт ионов через мембрану?
а) Потенциал покоя. На рисунке 1 показано, как осуществляется регистрация разности потенциалов с использованием одного электрода, размещенного на поверхности мембраны аксона, и второго электрода, подведенного к внутренней поверхности мембраны. В отсутствие стимуляции разность потенциалов составляет около 70 мВ. Несмотря на то что наружная поверхность мембраны на самом деле несет положительный заряд, ее заряд принято считать равным нулю. В покое потенциал на внутренней стороне мембраны составляет -70 мВ.
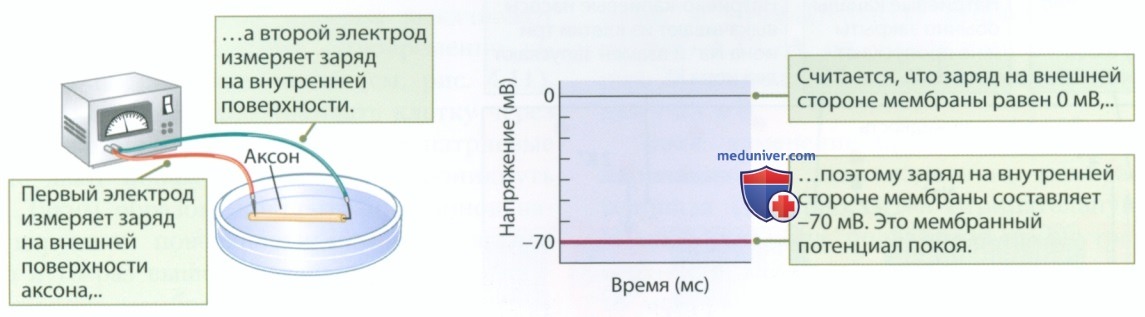
Почему ноль? Нам интересна разность потенциалов, а не собственно заряд.
Если бы мы продолжали запись в течение длительного времени, заряд в отсутствие стимуляции на мембране остался бы почти без изменений. Заряд может измениться в результате определенных изменений в мембране, однако в покое разница между зарядами на внешней и внутренней сторонах мембраны создает электрический потенциал, который дает возможность использовать накопленную энергию наподобие заряженной батарейки. Таким образом, заряд представляет собой запас энергии, называемый мембранным потенциалом покоя.
Термин «потенциал» в данном случае имеет примерно такое же значение, что и финансовый потенциал клиента банка — клиент копит деньги на будущее. Соответственно, потенциал покоя это запас энергии, который может быть использовать позднее. Потенциал покоя формируется в большинстве клеток организма, однако потенциалы покоя разных аксонов различаются. Значения потенциала покоя варьируют от -40 до -90 мВ в зависимости от типа нейрона и вида животного.
В формировании потенциала покоя принимают участие следующие виды заряженных частиц: ионы натрия (Na+), ионы калия (К+), ионы хлора (Cl-) и крупные отрицательно заряженные белковые молекулы (А-). Они представляют собой катионы и анионы, определение которых можно найти в отдельной статье на сайте - просим пользоваться формой поиска выше. Как показано на рис. 2, заряженные частицы распределены на мембране аксона неравномерно — внутриклеточная жидкость содержит больше белковых анионов и ионов калия, а во внеклеточной жидкости больше ионов натрия и хлора. С чем связана такая разница в концентрациях и как концентрация каждого из ионов влияет на потенциал покоя?
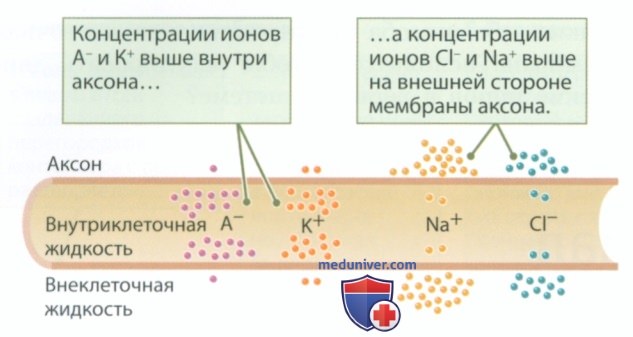
Чтобы запомнить, с какой стороны от мембраны накапливаются разные ионы, воспользуйтесь мнемонической фразой: поваренную соль (хлорид натрия) не сыплем в еду (то есть внутрь клетки), а ставим на стол отдельно (вне клетки).
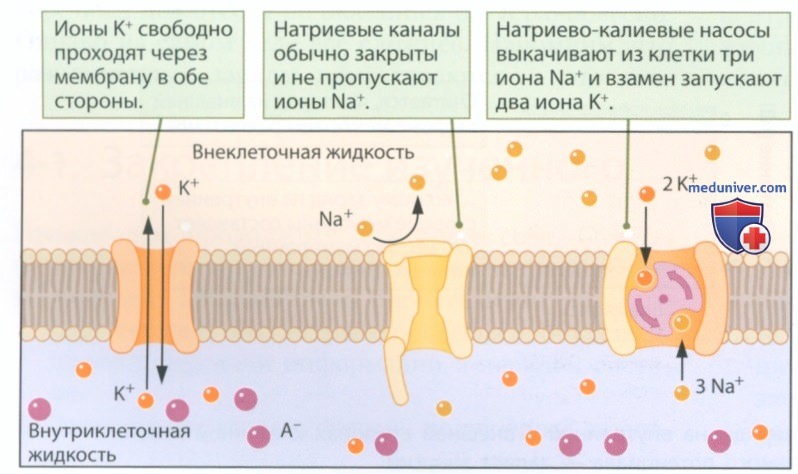
б) Поддержание потенциала покоя. За поддержание потенциала покоя отвечают ионные каналы, каналы с воротным механизмом и насосы клеточной мембраны. Рисунок 3, на котором изображена схема клеточной мембраны в покое, показывает, как перечисленные выше мембранные белки формируют заряд на поверхности мембраны в покое:
1) Поскольку мембрана практически непроницаема для крупных молекул, отрицательно заряженные белковые молекулы (А-) остаются внутри клетки.
2) Неуправляемые калиевые и хлорные каналы позволяют ионам калия (К+) и хлора (Cl-) свободно течь через мембрану, а воротный механизм натриевых каналов не пропускают положительно заряженные ионы натрия (Na+).
3) Натриево-калиевые насосы (Na+-K+) выкачивают из внутриклеточной жидкости ионы натрия (Na+) и взамен впускают в клетку ионы калия (К+).
1. Внутри клетки. Крупные белковые анионы образуются внутри клетки. Размеры мембранных каналов не позволяют этим белкам покинуть клетку, и их отрицательного заряда достаточно для формирования мембранного потенциала или потенциала покоя. Поскольку большинство клеток организма вырабатывает такие крупные отрицательно заряженные молекулы, на мембранах большей части клеток возникает разность потенциалов.
Чтобы уравновесить отрицательный заряд, создаваемый находящимися во внутриклеточной жидкости белковыми анионами, клетки накапливают положительно заряженные ионы калия — внутри клетки скапливается примерно в 20 раз больше ионов калия, чем снаружи от клетки. Ионы калия проходят через клеточную мембрану по открытым калиевым каналам, как показано на рис. 3. Однако при такой высокой концентрации ионов калия внутри клетки на мембране создается градиент концентрации ионов калия, который ограничивает число поступающих в клетку ионов калия. Другими словами, в клетку проникают не все ионы калия, которые могли бы в нее проникнуть.
Поскольку концентрация ионов калия внутри клетки значительно выше их концентрации снаружи, ионы калия выходят из клетки по градиенту концентрации.
Оставшихся снаружи от мембраны немногочисленных ионов калия достаточно для формирования мембранного потенциала. Они вносят свой вклад в результирующий отрицательный заряд на внутренней стороне мембраны. Последнее предложение может показаться странным. Если внутри клетки в 20 раз больше ионов калия, чем снаружи, почему же на внутренней стороне мембраны возникает отрицательный заряд? Разве все эти многочисленные ионы калия не должны создавать положительный заряд? Нет, потому что проникающих в клетку ионов калия недостаточно, чтобы уравновесить отрицательный заряд белковых анионов.
Давайте попробуем сформулировать так: если бы на внутренней стороне мембраны могло накапливаться неограниченное количество ионов калия, положительный заряд ионов калия точно соответствовал бы отрицательному заряду вырабатываемых внутри клетки белковых анионов и мембранный потенциал не возникал бы вообще. Однако количество проникающих в клетку ионов калия ограниченно: когда концентрация ионов К+ во внутриклеточной жидкости становится выше их концентрации вокруг клетки, дальнейший приток ионов калия становится невозможным из-за возникшего градиента концентрации.
2. Окружение клетки. Когда потенциал на мембране достигает равновесной величины по калию, часть ионов калия остается снаружи от клетки. Для поддержания отрицательного заряда на внутренней стороне мембраны снаружи от клетки должно быть совсем немного положительно заряженных ионов калия. Это значит, что ионы натрия вносят свой вклад в мембранный потенциал.
Ионы натрия (Na+) и хлора (Cl-) также участвуют в формировании потенциала покоя. Если бы положительно заряженные ионы натрия могли свободно проникать сквозь мембрану, они бы диффундировали в клетку. Это привело бы к исчезновению мембранного потенциала, возникающего в результате неравномерного распределения ионов натрия на внешней и внутренней сторонах мембраны. Такая диффузия не происходит, поскольку ворота натриевых каналов обычно закрыты (см. рис. 3) и большая часть ионов натрия не может попасть в клетку.
Тем не менее со временем в клетке могут накопиться ионы натрия, которых будет достаточно, чтобы нейтрализовать мембранный потенциал. Клеточная мембрана использует специфический механизм, чтобы не допустить нейтрализации мембранного потенциала.
Когда ионы натрия проникают внутрь нейрона, их немедленно выбрасывают из клетки натри-ево-калиевые насосы — встроенные в клеточную мембрану белковые молекулы. Мембрана содержит многие тысячи насосов, которые выкачивают из клетки по три иона натрия, одновременно запуская внутрь по два иона калия (см. рис. 3). Ионы калия могут свободно покидать клетку через открытые калиевые каналы, а закрытые натриевые каналы не дают ионам натрия снова проникнуть в клетку. Таким образом, концентрация ионов натрия на наружной поверхности мембраны аксона примерно в 10 раз выше их концентрации на внутренней стороне мембраны. Разница в концентрациях ионов натрия также вносит свой вклад в потенциал покоя.
Теперь рассмотрим ионы хлора. В отличие от ионов натрия, ионы хлора могут свободно течь через мембрану в обоих направлениях. Равновесная величина, когда градиент концентрации становится равным градиенту напряжения, примерно равна мембранному потенциалу покоя, поэтому вклад ионов хлора в потенциал покоя обычно невелик. Равновесие по хлору обеспечивает концентрация ионов хлора вокруг клетки, примерно в 12 раз превышающая их концентрацию внутри клетки.
Избирательная проницаемость клеточной мембраны и действие ионных каналов, каналов с воротным механизм и насосов способствуют формированию мембранного потенциала — потенциала покоя (рис. 4).

в) Градуальные потенциалы. Потенциал действия обеспечивает запас энергии, который используется примерно как вода в плотине — водяные ворота открываются, выпуская воду, необходимую для орошения или производства электроэнергии. Если в отсутствие стимуляции концентрация любого из ионов на клеточной мембране меняется, то меняется мембранный потенциал. Градуальные потенциалы — это небольшие изменения мембранного потенциала.
Электростимуляция мембраны с помощью микроэлектрода позволяет воспроизвести процесс изменения мембранного потенциала и формирования градуального потенциала в живой клетке. Если подаваемое на внутреннюю поверхность мембраны напряжение отрицательное, мембранный потенциал меняется на несколько милливольт в отрицательную сторону. Как показано на рис. 5, он может измениться по сравнению с равным -70 мВ потенциалом покоя, составив -73 мВ.
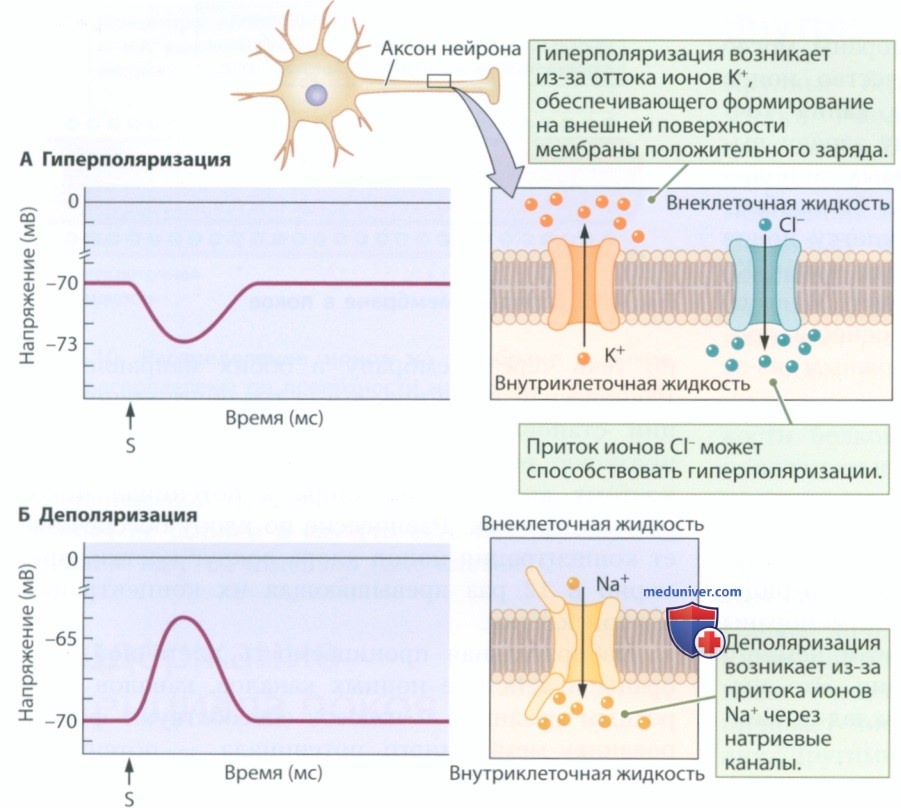
Такое изменение представляет собой гиперполяризацию, то есть изменение мембранного потенциала (полярности) в отрицательную сторону. И наоборот, если подаваемое на внутреннюю поверхность мембраны напряжение положительное, то мембранный потенциал меняется на несколько милливольт в положительную сторону. Как показано на рис. 5, он может немного отклониться от равного -70 мВ потенциала покоя, составив -65 мВ. Такое изменение представляет собой деполяризацию, то есть изменение мембранного потенциала в положительную сторону. Градуальные потенциалы обычно длятся лишь несколько миллисекунд.
Гиперполяризация и деполяризация обычно возникают на мембране в области тела клетки (сомы) и дендритов; там находятся каналы с воротным механизмом, которые открываются и закрываются, изменяя мембранный потенциал (см. рис. 5).
Градуальные потенциалы возникают благодаря функционированию калиевых, хлорных и натриевых каналов:
1. Калиевые каналы. Для гиперполяризации требуется сдвиг заряда на внешней стороне мембраны в положительную сторону — для этого необходим усиленный отток ионов калия. Если калиевые каналы обычно находятся в открытом состоянии, за счет чего может произойти усиление оттока ионов калия? Судя по всему, даже при открытых калиевых каналах ряд факторов препятствует оттоку ионов калия. Снижение влияния этих факторов приводит к гиперполяризации.
2. Хлорные каналы. Гиперполяризация мембраны также может возникнуть вследствие притока ионов хлора. Несмотря на то что ионы хлора могут свободно течь через мембрану, большинство ионов хлора остается снаружи. Снижение влияния факторов, препятствующих току ионов хлора, может приводить к кратковременному повышению уровня Cl- в клетке.
3. Натриевые каналы. Деполяризация может возникать в тех случаях, когда обычно закрытые ворота натриевого канала открываются, обеспечивая приток ионов натрия.
Доказательством участия ионов калия в гиперполяризации является способность блокатора калиевых каналов тетраэтиламмония блокировать гиперполяризацию. Об участии натриевых каналов в деполяризации говорит способность блокатора натриевых каналов тетродотоксина блокировать деполяризацию. Рыба фугу, которую в ряде стран, в том числе в Японии, считают деликатесом, содержит тетродотоксин, необходимый ей для защиты от хищников. Чтобы приготовить такую рыбу, нужно обладать особыми навыками. Употребление неправильно приготовленной рыбы может привести к летальному исходу, потому что содержащийся в ней яд подавляет электрическую активность нейронов.
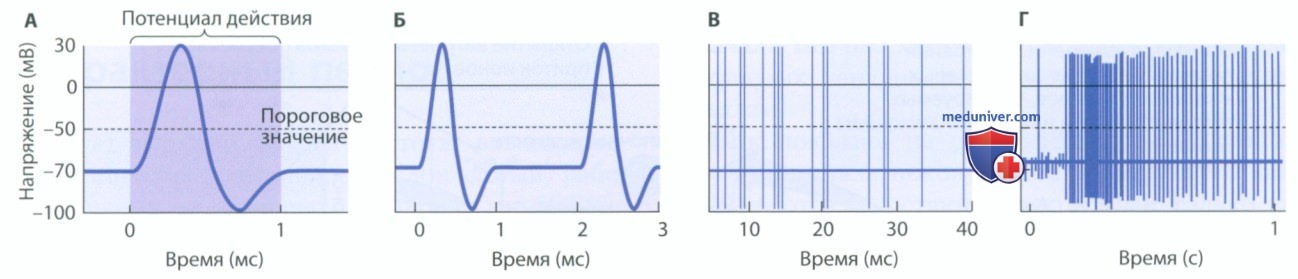
г) Потенциал действия. Электростимуляция находящейся в покое клеточной мембраны обеспечивает формирование локальных градуальных потенциалов. В то же время, потенциал действия — это кратковременное (около 1 мс), но значительное изменение полярности мембраны (рис. 6). Мембранный потенциал внезапно меняется таким образом, что внутренняя поверхность мембраны становится положительно заряженной по отношению к внешней поверхности, а затем меняется обратно, обеспечивая восстановление потенциала покоя. Поскольку длительность потенциала действия крайне мала, в течение секунды может возникнуть множество потенциалов действия — это можно увидеть на рис. 6, где представлены сжатые временные шкалы.
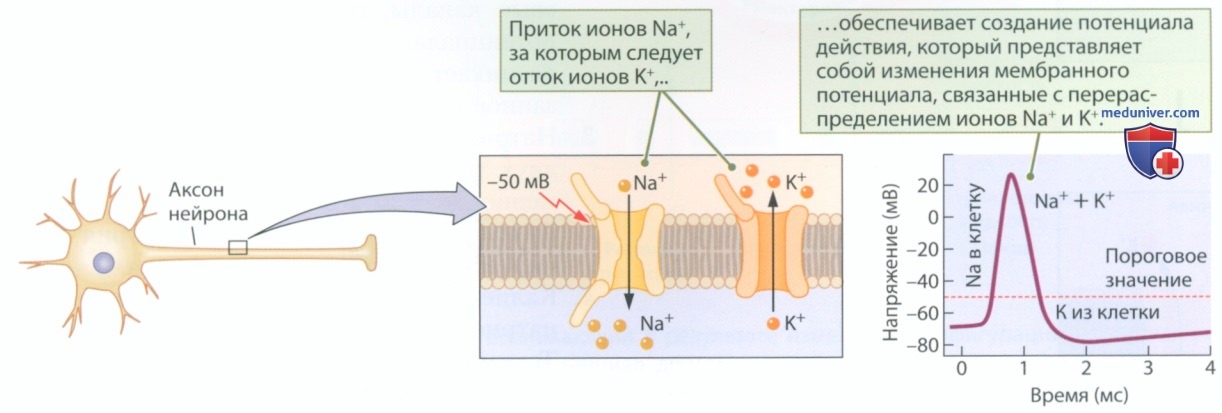
Потенциал действия возникает, когда мембрану в течение короткого периода пересекает большое количество ионов Na+ и К+. Фаза деполяризации соответствует притоку ионов Na+, а фаза гиперполяризации соответствует оттоку ионов К+. Ионы натрия устремляются в клетку, за этим следует отток ионов калия из клетки. Как показано на рис. 7, в формировании потенциала действия ведущую роль играют ионы натрия и калия.
Потенциал действия возникает при деполяризации клеточной мембраны примерно до -50 мВ. Перешагнув через указанное пороговое значение, мембранный потенциал претерпевает значительные изменения без дополнительной стимуляции. Мембранный потенциал повышается до нуля, обеспечивая дальнейшую деполяризацию. Процесс деполяризации продолжается до тех пор, пока заряд на внутренней стороне мембраны не достигнет +30 мВ. Таким образом, сдвиг мембранного потенциала составляет 100 мВ. Затем мембранный потенциал снова меняется, обеспечивая небольшую гиперполяризацию. При этом сдвиг мембранного потенциала немного превышает 100 мВ. После этого мембрана постепенно восстанавливает потенциал покоя, который составляет -70 мВ.
В норме потенциал действия создают изменения, вызванные сначала притоком ионов натрия, а затем оттоком ионов калия из аксона. Результаты эксперимента показали, что электростимуляция мембраны аксона, который поместили в раствор тетраэтиламмония (блокатор калиевых каналов), способствует уменьшению потока ионов по сравнению с нормой за счет притока ионов Na+. Аналогичным образом стимуляция мембраны аксона, находящегося в растворе тетродотоксина (блокатор натриевых каналов), приводит к изменению потока ионов за счет оттока ионов К+. Результаты эксперимента показаны на рис. 8, который представляет собой схему изменений потока ионов, а не мембранного потенциала.
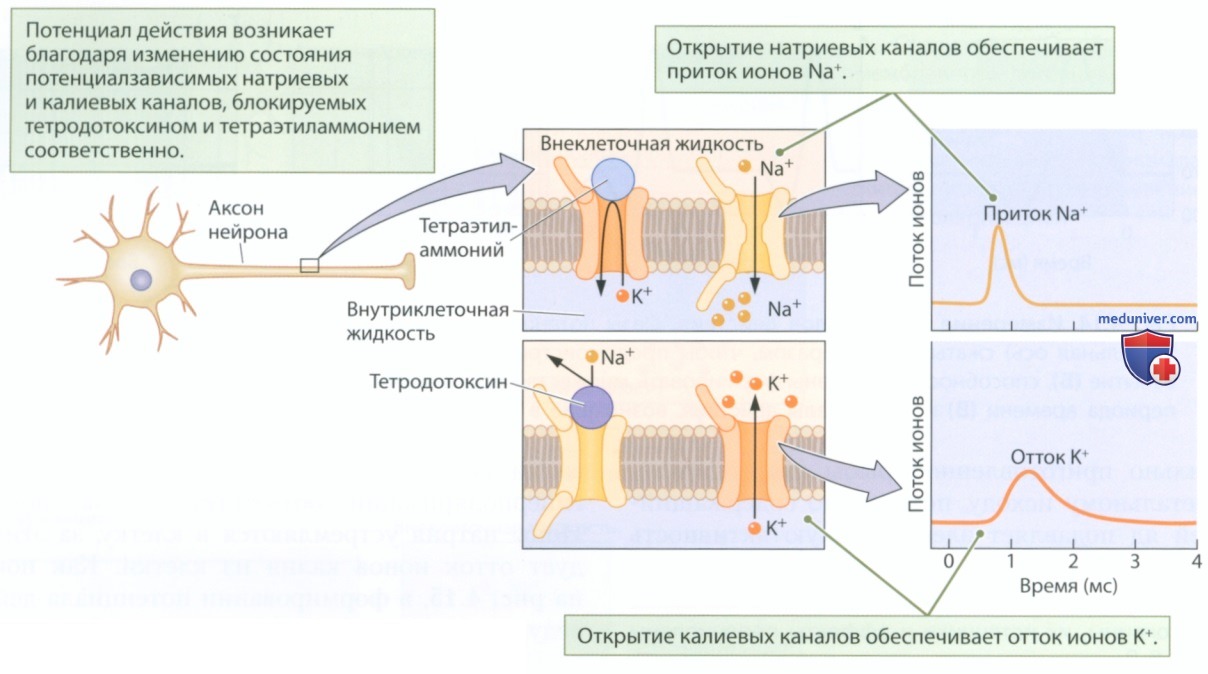
1. Роль потенциалзависимых ионных каналов. Какие клеточные механизмы обеспечивают транспорт ионов натрия и калия, генерируя потенциал действия? Это натриевые и калиевые каналы с воротным механизмом, которые открываются и закрываются в ответ на изменение мембранного потенциала (рис. 9).
Такие потенциалзависимые каналы остаются закрытыми, пока мембрана аксона находится в покое (потенциал покоя) — ионы не могут пройти через них. Когда мембранный потенциал достигает порогового значения, конфигурация потенциалзависимых ионных каналов меняется — они ненадолго открываются, пропуская ионы, а затем снова закрываются, препятствуя транспорту ионов. Имеет место следующая последовательность событий:
1) Потенциалзависимые натриевые и калиевые каналы реагируют на пороговый потенциал, величина которого составляет около -50 мВ. Когда мембранный потенциал достигает такой величины, каналы обоих типов открываются, позволяя ионам течь через мембрану.
2) Потенциалзависимые натриевые каналы активируются быстрее, чем потенциалзависимые калиевые каналы, поэтому изменение мембранного потенциала, связанное с притоком ионов Na+, возникает немного раньше, чем изменение, связанное с оттоком ионов К+.
3) Натриевые каналы имеют воротный механизм двух типов. При деполяризации мембраны примерно до +30 мВ первые ворота закрываются. Таким образом, приток ионов натрия быстро начинается и быстро заканчивается.
4) Калиевые каналы открываются медленнее, чем натриевые, и дольше остаются открытыми. Таким образом, отток ионов К+ устраняет вызванную притоком ионов Na+ деполяризацию, приводя к небольшой гиперполяризации мембраны.
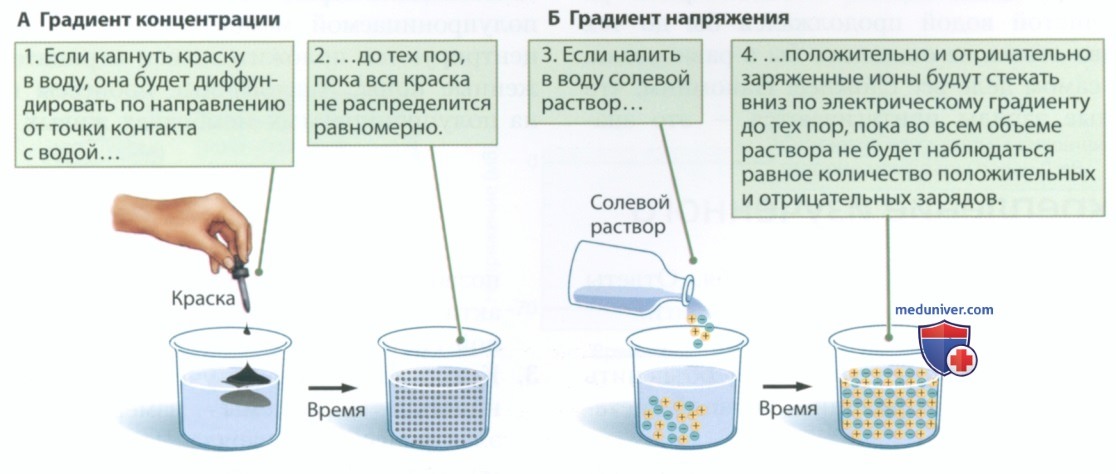
2. Потенциал действия и рефрактерный период. Существует верхний предел частоты возникновения потенциалов действия, обусловленный работой натриевых и калиевых каналов. Стимуляция мембраны аксона в фазе деполяризации потенциала действия не приводит к формированию нового потенциала действия. Находясь в фазе реполяризации, аксон также не способен создать потенциал действия. В этот период мембрана находится в состоянии абсолютной рефрактерности.
P.S. Есть исключения: некоторые нейроны ЦНС разряжаются в фазе реполяризации.
Тем не менее стимуляция мембраны аксона в фазе гиперполяризации может индуцировать новый потенциал действия, однако повторная стимуляция должна быть более интенсивной, чем первоначальная. В этой фазе мембрана находится в состоянии относительной рефрактерности.
Рефрактерный период определяет поведение потенциалзависимых натриевых и калиевых каналов. Натриевый канал содержит ворота двух типов, а калиевый канал — ворота одного типа. На рисунке 10 показано положение ворот, соответствующее различным фазам потенциала действия. Сначала мы рассмотрим изменения конфигурации натриевых каналов, а затем перейдем к калиевым каналам.
Как показано на рис. 10, после восстановления потенциала покоя ворота 1 натриевого канала закрываются — остаются открытыми только ворота 2. Когда стимул достигает порогового значения, ворота 1 тоже открываются, после чего быстро закрываются ворота 2. Такая последовательность событий обеспечивает короткий период, в течение которого остаются открытыми ворота обоих типов. Пока открыты ворота обоих типов и после закрытия ворот 2 мембрана находится в состоянии абсолютной рефрактерности.
Открытие калиевых каналов приводит к реполяризации и последующей гиперполяризации клеточной мембраны. Калиевые каналы открываются и закрываются медленнее, чем натриевые. Гиперполяризация, вызванная продолжающимся оттоком ионов калия, затрудняет деполяризацию мембраны до порогового значения, которое вновь открывает ворота, задействованные в формировании потенциала действия. На время гиперполяризации мембрана находится в состоянии относительной рефрактерности.
Изменения полярности, связанные с формированием потенциала действия, можно сравнить с работой унитаза, снабженного рычагом для слива воды. При слабом нажатии на рычаг сливается небольшое количество воды; подача воды прекращается, когда вы отпускаете рычаг. Это похоже на градуальный потенциал. Более сильное нажатие на рычаг запускает смыв, при этом подаваемый объем воды непропорционален силе нажатия на рычаг. Это похоже на потенциал действия.
Пока идет смыв, унитаз находится в состоянии абсолютной рефрактерности — повторное нажатие на рычаг не позволяет запустить смыв еще раз. Пока идет заполнение бачка, унитаз находится в состоянии относительной рефрактерности — запустить смыв можно, но для этого придется сильнее нажать на рычаг. Полноценный смыв можно будет запустить только после того, как цикл завершится и унитаз вернется в состояние покоя.
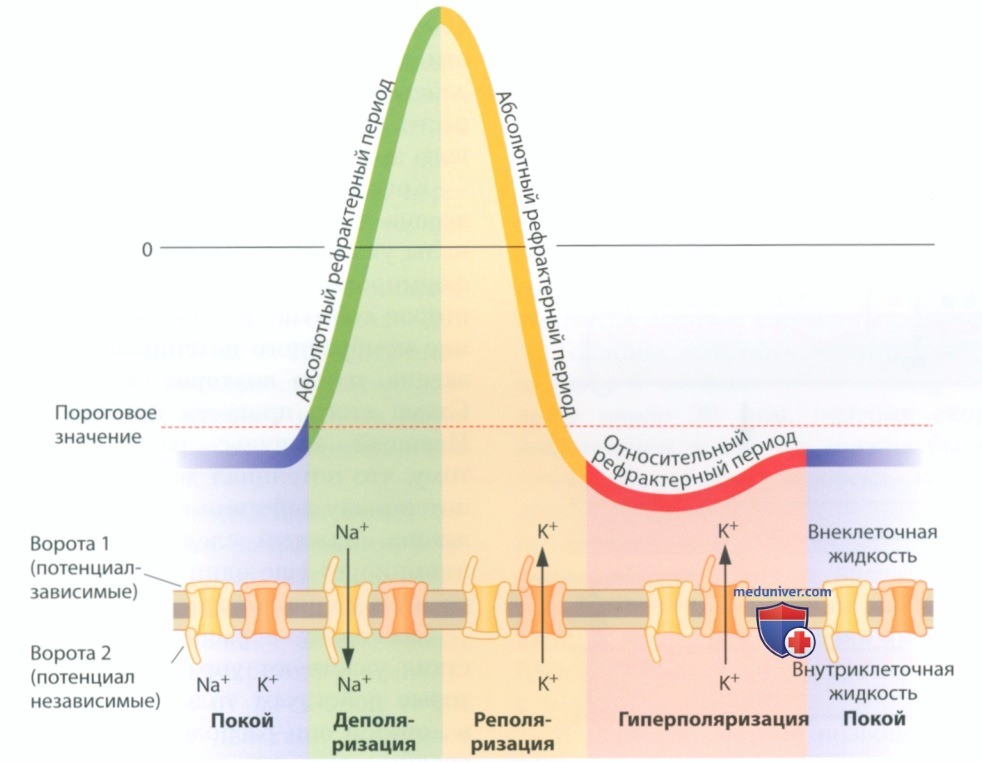
д) Нервный импульс. Предположим, что мы разместили два регистрирующих электрода на мембране аксона, установив их на некотором расстоянии друг от друга, а затем начали стимулировать область рядом с одним из электродов. Этот электрод немедленно зарегистрирует потенциал действия. Второй электрод сразу же зарегистрирует такой же сигнал. Рядом со вторым электродом тоже возникает потенциал действия, несмотря на то что этот электрод установлен на некотором расстоянии от точки стимуляции.
Может быть второй потенциал действия — всего лишь распространяющееся вдоль аксона эхо первого потенциала действия? Нет, это не так — размер и форма зарегистрированных двумя электродами сигналов совершенно идентичны. Второй потенциал действия не является ослабленной версией первого — он равен ему по величине. Каким-то образом потенциал действия перемещается вдоль аксона. Такое распространение потенциала действия по аксону называют нервным импульсом.
Почему распространяется потенциал действия? Напомним, что сопровождающий потенциал действия сдвиг мембранного потенциала составляет 100 мВ — он намного превышает сдвиг в 20 мВ, необходимый для перехода мембраны из состояния покоя (-70 мВ) к пороговому потенциалу -50 мВ. Следовательно, изменение напряжения на той части мембраны, где впервые возникает потенциал действия, достаточно велико — оно позволяет довести близлежащие области мембраны до порогового значения в -50 мВ.
Когда мембранный потенциал соседней области аксона достигает -50 мВ, потенциалзависимые каналы указанной области открываются, обеспечивая формирование в ней потенциала действия. Это второе событие, в свою очередь, вызывает изменение мембранного потенциала в следующей области аксона, и это повторяется по всей длине аксона. Схема этого процесса представлена на рис. 11. Нервный импульс распространяется благодаря тому, что потенциал действия дает начало другому потенциалу действия в соседней области мембраны аксона.
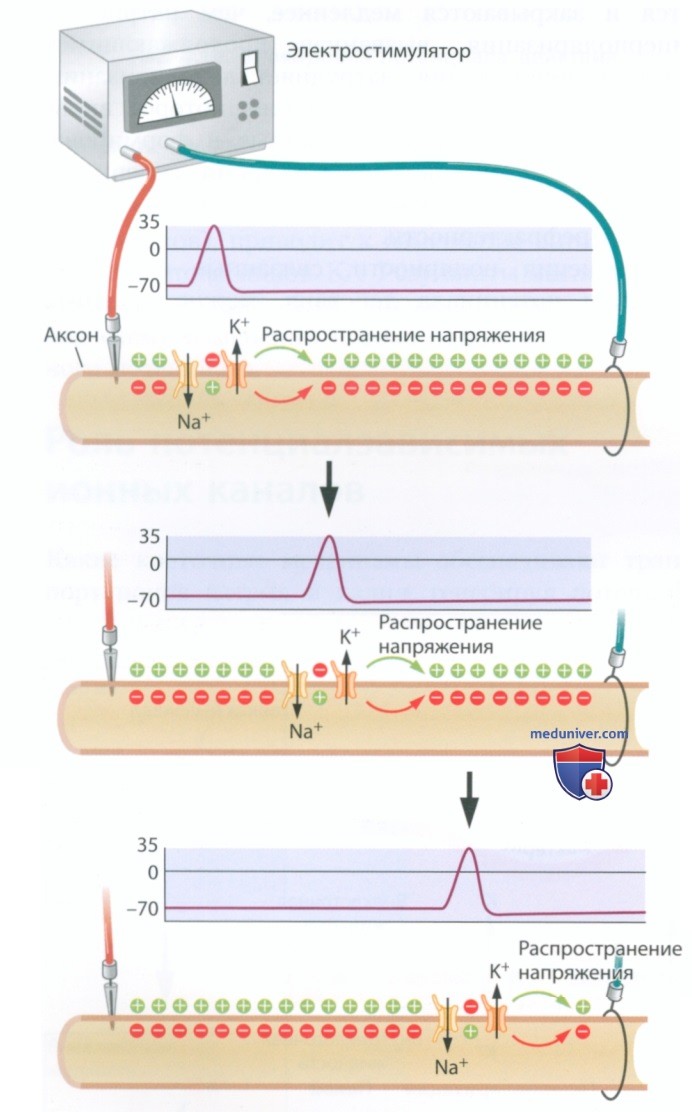
Каждый следующий потенциал действия инициирует еще один потенциал действия — и так по всей длине нервного волокна.
Поскольку, в распространении потенциала действия участвуют управляемые ионные каналы, которые действуют только на ту область мембраны, в которой они расположены, потенциалы действия, возникающие по всей длине нерва или тракта, имеют одинаковую величину. Потенциал действия развивается за счет потенциальной энергии определенной области мембраны, и такое же количество энергии обычно расходуется на каждом участке мембраны при распространении нервного импульса.
В итоге потенциалы действия не рассеиваются — они либо возникают в полном объеме, либо не возникают вообще, подчиняясь закону «все или ничего». Потенциал действия, формирующий нервный импульс или сообщение, имеет постоянные характеристики — он передается на каждую терминаль нерва в неизмененном виде.
Давайте представим себе расположенные вдоль аксона потенциалзависимые каналы в виде костяшек домино. Когда одна из них падает, она толкает соседнюю костяшку — и так по всей линии. Падают абсолютно все костяшки. Последняя костяшка преодолевает такое же расстояние и падает так же сильно, как и первая.

Фактически эффект домино возникает, когда открываются потенциалзависимые каналы. Открытие одного канала создает сдвиг мембранного потенциала, заставляющий открыться соседний канал — точно так же одна костяшка домино толкает соседнюю костяшку, заставляя ее упасть. Каналы открываются по всей длине аксона, и последний канал открывается точно так же, как и первый — точно так же падает вся длинная цепочка домино.
е) Рефрактерные периоды и функционирование нервных волокон. Рефрактерные периоды определяет положение управляющих потоком ионов ворот потенциалзависимых каналов. Из-за этого максимальная частота возникновения потенциалов действия — это примерно один потенциал действия в 5 мс. При передаче информации нервы используют рефрактерные периоды двумя способами.
Во-первых, максимальная частота возникновения потенциалов действия составляет около 200 в секунду (1с, или 1000 мс/5 мс = 200 потенциалов действия за 1 с). Чувствительность потенциалзависимых ионных каналов, разная у разных типов нейронов, также влияет на частоту возникновения потенциалов действия.
Во-вторых, хотя потенциал действия может перемещаться по аксону в любом направлении, рефрактерные периоды не позволяют ему изменить направление и вернуться в исходную точку. Таким образом, рефрактерные периоды обеспечивают формирование единичного дискретного импульса, который распространяется от начальной точки стимуляции. Если потенциал действия возникает в области тела клетки, он обычно распространяется по аксону до терминалей.
Давайте еще раз вспомним эффект домино: если костяшки упали, нужно время, чтобы заново поднять их. Это и есть рефрактерный период. Поскольку каждая костяшка падает, толкая соседнюю, последовательность не изменится, пока костяшки не будут снова установлены вертикально: они могут падать только в одном направлении.
Тот же принцип определяет направление распространения потенциала действия.
ж) Сальтаторное проведение и миелиновая оболочка. Крупные размеры гигантских аксонов кальмара обеспечивают очень быструю передачу нервных импульсов — точно так же труба большого диаметра быстро пропускает через себя большой объем воды. Однако крупные аксоны занимают немало места -у кальмара не может быть большого количества таких аксонов, иначе его тело стало бы слишком громоздким. У млекопитающих, в организме которых присутствует множество иннервирующих мышцы аксонов, они не могут быть крупными. Наши аксоны должны быть как можно более тонкими, потому что для производства сложных движений нам требуется великое множество аксонов.
Толщина наиболее крупных аксонов человека, идущих к мышцам и от мышц, составляет всего лишь около 30 мкм, поэтому скорость передачи информации в них не должна быть особенно высокой. И все-таки человека, как и большинство других млекопитающих, нельзя назвать медлительным существом. Мы обрабатываем информацию и генерируем ответ с впечатляющей скоростью. Как нам это удается, если наши аксоны такие тонкие? В процессе эволюции позвоночных эта проблема была решена, и ее решение никак не связано с толщиной аксонов.
В нервной системе позвоночных ускорение передачи нервных импульсов обеспечивают глиальные клетки. Шванновские клетки периферической нервной системы и олигодендроциты центральной нервной системы человека оборачиваются вокруг аксонов, образуя миелиновую оболочку (рис. 12). Потенциалы действия не могут возникать в местах, где аксон покрыт миелиновой оболочкой. Во-первых, миелин выполняет функцию барьера, препятствующего потоку ионов. Во-вторых, покрытые миелином области аксона содержат мало ионных каналов, необходимых для создания потенциала действия.
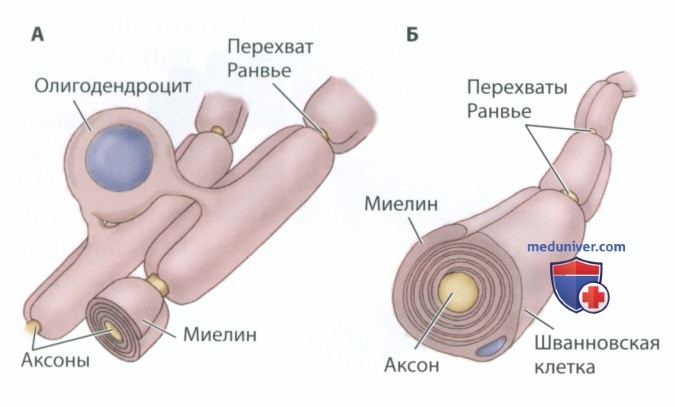
Однако не весь аксон покрыт миелином. Зазоры между миелинизирующими глиальными клетками богаты потенциалзависимыми ионными каналами. Такие разрывы в миелиновой оболочке, называемые перехватами Ранвье, располагаются довольно близко друг к другу, и потенциал действия, возникший на одном перехвате Ранвье, может открыть потенциалзависимые ворота на соседнем перехвате. Таким образом, сравнительно медленный потенциал действия может быстро «перескакивать» с одного перехвата Ранвье на другой (рис. 13). Такое проведение нервного импульса назвали сальтаторным проведением (от лат. saltare — скакать, прыгать).
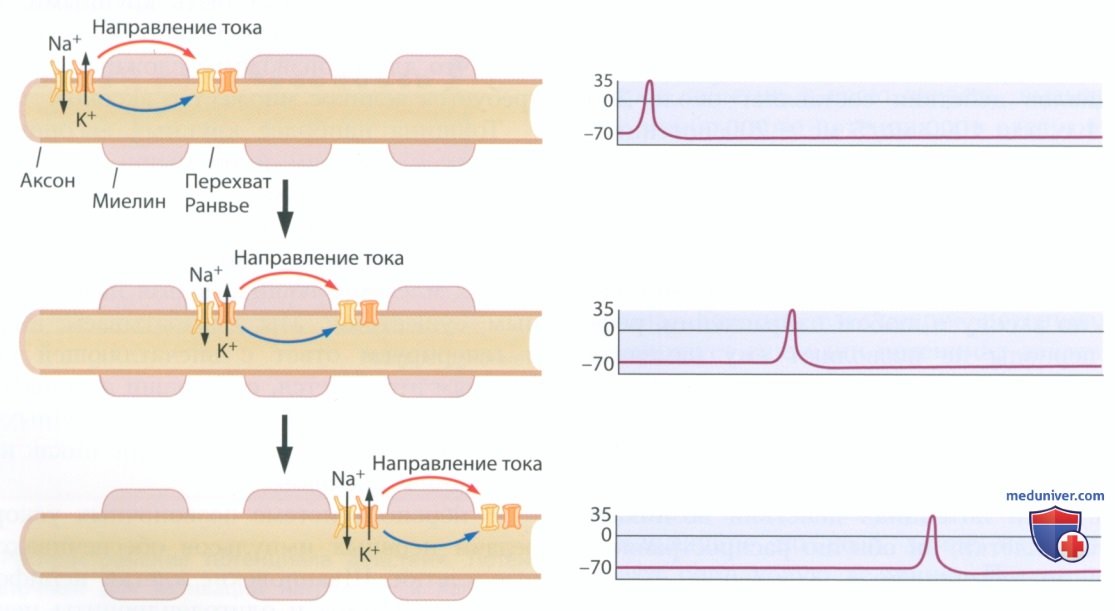
Миелин обеспечивает два важных преимущества при распространении потенциалов действия. Во-первых, распространение потенциала действия становится менее энергоемким процессом, поскольку потенциалы действия возникают только на перехватах Ранвье, а не по всей длине аксона. При этом для распространения потенциала действия в иемиелинизированных аксонах требуется значительный запас метаболической энергии (Crotty et al., 2006). Второе преимущество состоит в том, что миелин ускоряет распространение потенциала действия.
Перескакивание с одного перехвата Ранвье на другой позволяет потенциалу действия быстрее распространяться по аксону, потому что электрический ток, текущий в аксоне под миелиновой оболочкой, распространяется очень быстро. Если ток движется быстро, то напряжение постепенно падает по мере продвижения по участку. Перехваты Ранвье расположены идеально: они обеспечивают достаточное напряжение на следующем перехвате, позволяющее преодолеть пороговый потенциал и, таким образом, восстановить потенциал действия.
В крупных миелинизированных аксонах млекопитающих нервные импульсы распространяются со скоростью до 120 м/с. В более мелких немиелинизированных аксонах скорость передачи нервного импульса составляет около 30 м/с.
Болельщики на спортивных матчах иногда создают волну, которая обходит вокруг стадиона. В тот момент, когда один человек встает, второй начинает вставать, создавая эффект волны. С такой волной можно сравнить проведение сигнала немиелинизированными аксонами. Теперь давайте представим себе, насколько быстрее волна обойдет вокруг стадиона, если встанут только зрители, занимающие угловые места. То же самое происходит с нервным импульсом, который распространяется, перескакивая с одного перехвата Ранвье на другой. Сальтаторное проведение отчасти определяет быстроту реакций человека и других млекопитающих.
Нейроны, которые должны быстро передавать информацию на большое расстояние, в том числе чувствительные нейроны и мотонейроны, имеют толстую миелиновую оболочку. Если миелин поврежден, нейрон может утратить способность передавать информацию. При рассеянном склерозе (PC) поражаются образованные олигодендроцитами миелиновые оболочки аксонов, что приводит к нарушению функции нейронов. Клиническая картина рассеянного склероза подробно описана в разд. «Клинические аспекты 4-2: Рассеянный склероз».
з) Клинические аспекты. Рассеянный склероз. Однажды Дж. О., которая только что окончила университет и начала работать бухгалтером, обратила внимание на помутнение в правом глазу. Она потерла глаз, но помутнение не исчезло. Более того, через несколько дней оно увеличилось в размерах.
Офтальмолог порекомендовал Дж. О. обратиться к неврологу, который диагностировал у нее неврит зрительного нерва, который может быть одним из симптомов рассеянного склероза (PC).
PC возникает в результате разрушения образованных олигодендроцитами миелиновых оболочек в ЦНС (см. рис. ниже). Пораженные нейроны теряют способность распространять потенциалы действия посредством сальтаторного проведения. Разрушение миелина носит очаговый характер — в пораженных областях возникают твердые рубцы. (Слово «склероз» в переводе с греч. означает «рубец».)
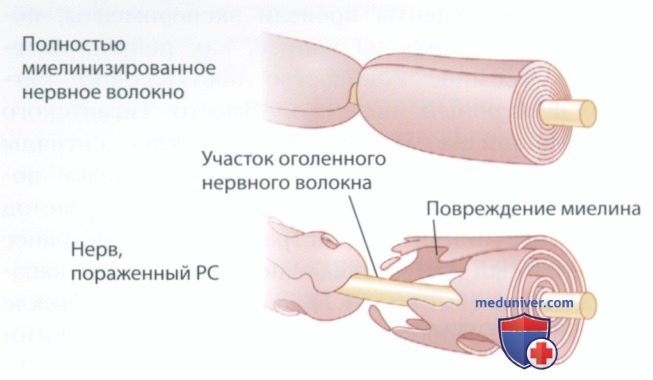
На пораженном участке образуется очаг разрушения миелина или твердая бляшка, которую можно визуализировать методом магнитно-резонансной томографии (МРТ). Разрушение миелина приводит к нарушению функции нейронов, с которым связаны характерные симптомы PC, такие как нарушения чувствительности и двигательные нарушения.
PC обычно сопровождается утомляемостью, болью и депрессией. Осложнениями PC являются дисфункция мочевого пузыря, запор и сексуальная дисфункция. PC в два раза чаще встречается у женщин, чем у мужчин. Заболевание оказывает значительное влияние на эмоциональную, социальную и профессиональную жизнь больного.
Рассеянный склероз — самое распространенное из примерно 80 аутоиммунных заболеваний, при которых иммунная система вырабатывает антитела, атакующие здоровые ткани организма (Rezania et al., 2012). Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) одобрило 15 препаратов, изменяющих течение рассеянного склероза, однако полностью излечить заболевание с помощью существующих методов вряд ли возможно (Reich et al., 2018).
Через несколько месяцев помутнение в правом глазу Дж. О. исчезло. Новые симптомы не появлялись еще три года — до рождения первого ребенка. После родов Дж. О. почувствовала покалывание в правой руке. Покалывание распространилось по всей руке, после чего рука Дж. О. на пять месяцев утратила подвижность. Затем подвижность в руке восстановилась. Однако пять лет спустя, после рождения второго ребенка, Дж. О. почувствовала покалывание в большом пальце левой ноги, которое впоследствии распространилось по всей ноге и снова привело к утрате подвижности.
Дж. О. лечили кортикостероидами, которые помогали, однако после отмены препаратов симптомы возвращались. Потом симптомы стали слабеть и в конце концов исчезли.
С тех пор у Дж. О. больше не возникали серьезные двигательные нарушения. Однако она жаловалась на сильную утомляемость, долго спала днем и рано ложилась спать по вечерам. У сестры и двоюродной сестры Дж. О. были похожие симптомы. У третьей сестры те же симптомы возникли в среднем возрасте, а у дочери Дж. О. — в возрасте около 25 лет. Одна из бабушек Дж. О. была прикована к инвалидному креслу, однако причина ее болезни неизвестна.
PC трудно диагностировать. Симптомы обычно появляются в зрелом возрасте. Заболевание проявляется внезапно, быстро приводя к тяжелым последствиям. Первые симптомы — это утрата чувствительности в лице, конечностях или теле, двигательные нарушения, а также сочетание нарушений чувствительности и двигательных нарушений. Двигательные симптомы чаще проявляются в руках или ногах.
За первым проявлением заболевания часто следует ремиссия, которая может длиться годами. Тем не менее некоторые формы PC прогрессируют очень быстро и пациент уже через несколько лет оказывается прикованным к постели.
PC чаще встречается в самых северных и самых южных широтах, что говорит о его возможной связи с дефицитом витамина D, вырабатываемого при воздействии солнечного света на кожу. Заболевание также может возникнуть из-за генетической предрасположенности — как и в случае с Дж. О. Многие пациенты с PC принимают витамины D3 и В12.
Хотя PC является самым распространенным хроническим воспалительным заболеванием ЦНС, которым страдают более 2 млн человек по всему миру (Reich et al., 2018), причины воспаления и разрушения миелиновых оболочек по-прежнему не выяснены. Ученые предположили, что рассеянный склероз в первую очередь является дегенеративным заболеванием, вызывающим вторичную аутоиммунную реакцию (Stys, 2013). Исследования этого вопроса очень важны, потому что ответ на него позволит изменить подход к лечению данного заболевания.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) - профессор, д.м.н. П.Е. Умрюхин
- Читать далее "Как нейроны интегрируют информацию: суммация, универсальность нейрона"
Редактор: Искандер Милевски. Дата публикации: 12.7.2023
- История поиска источников электрической активности в нервной системе
- Мембранный потенциал: потенциал покоя, потенциал действия, нервный импульс, рефрактерный период
- Как нейроны интегрируют информацию: суммация, универсальность нейрона
- Доставка информации в нервную систему и обратно
- Резюме по использованию электрических сигналов нейронами для передачи информации