Как нейроны интегрируют информацию: суммация, универсальность нейрона
Дендриты нейрона покрыты множеством шипиков, позволяющих нейрону установить свыше 50 000 соединений с другими нейронами. Тело нейрона, лежащее между дендритами и аксоном, способно принимать большое количество сигналов. Нервные импульсы, передаваемые на нейрон через множество синаптических контактов, бомбардируют нейрон возбуждающими и тормозными сигналами.
В 1950-1960-х гг. Джон Экклс (John С. Eccles, 1965) и его студенты провели эксперименты, позволившие ответить на вопрос, как нейрон интегрирует огромное количество поступающих сигналов в нервный импульс. Вместо гигантского аксона кальмара Экклс регистрировал сигналы тел крупных мотонейронов спинного мозга позвоночных. Для этого он усовершенствовал метод электростимуляции и регистрации сигналов, ранее разработанный для исследования гигантских аксонов кальмара (см. рис. 4.1). За свою работу Экклс был удостоен Нобелевской премии по физиологии и медицине.
Например, мотонейроны аккумулируют сигналы из многих источников. Мотонейроны спинного мозга образуют множество дендритов, каждый из которых дает около 20 основных ветвлений, которые в свою очередь образуют ответвления. Поверхность дендрита усеяна дендритными ши-пиками. Способность принимать сигналы от кожи, суставов, мышц, спинного мозга и головного мозга делает мотонейроны идеальным объектом для изучения реакции нейронов на разнообразные сигналы. Аксон каждого мотонейрона иннервирует мышцу. Мотонейрон — это последняя инстанция, которую нервная система использует для формирования поведения.
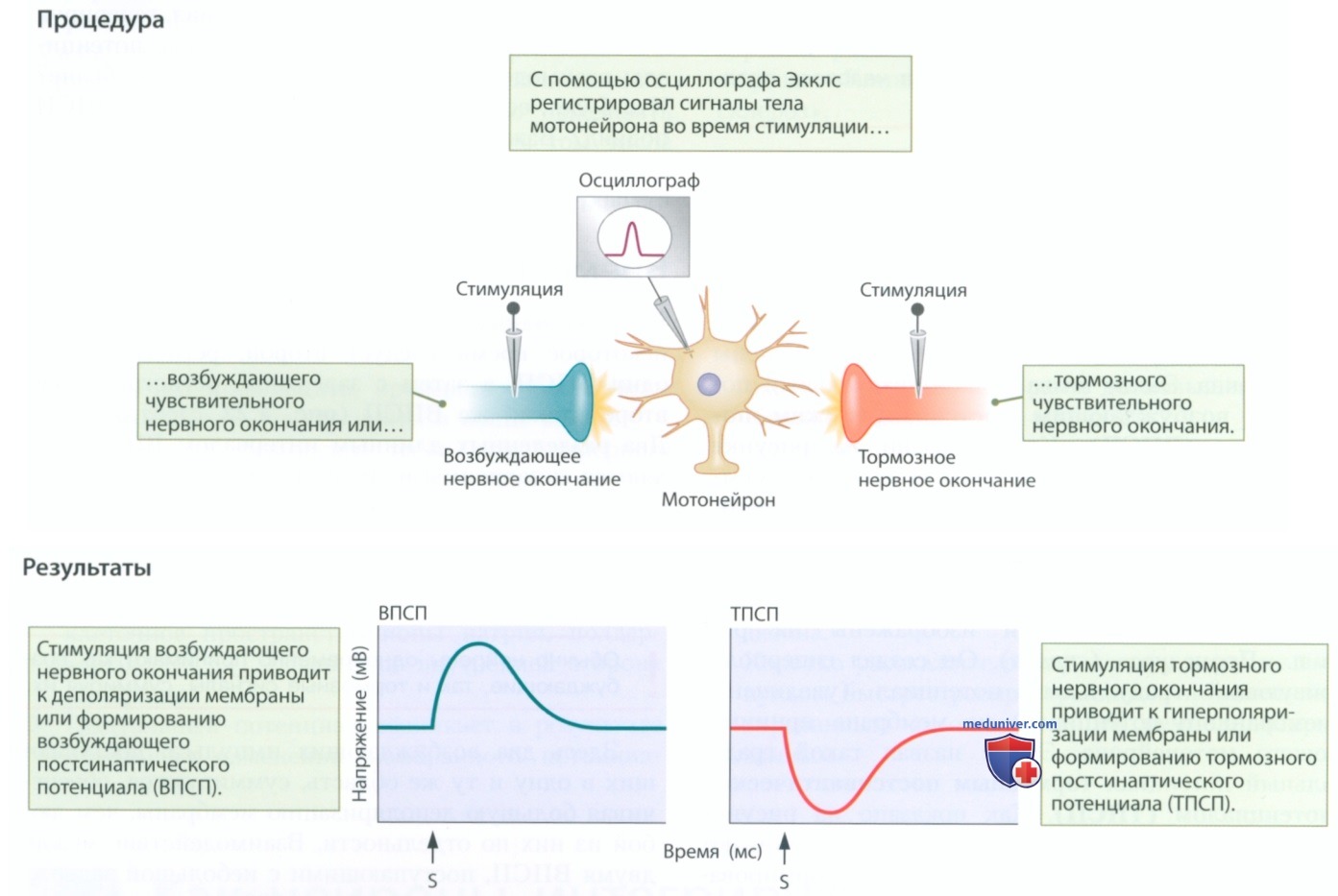
а) Возбуждающие и тормозные постсинаптические потенциалы. Чтобы исследовать активность мотонейрона, Экклс ввел микроэлектрод в спинной мозг позвоночного животного таким образом, чтобы кончик микроэлектрода оказался внутри тела мотонейрона или рядом с ним. Затем он установил стимулирующие электроды на аксоны идущего к спинному мозгу сенсорного нервного волокна. Разделив сенсорные нервные волокна, он получил возможность стимулировать нервные волокна по отдельности.
1. Эксперимент. Как стимуляция влияет на возбудимость нейрона?
Выводы. ВПСП повышает вероятность того, что результатом будет потенциал действия. ТПСП препятствуют формированию потенциала действия.
Описание проведенного Экклсом эксперимента. Как показано на рисунке в п. «Процедура», стимуляция некоторых сенсорных волокон обеспечивала формирование деполяризующего градуального потенциала (уменьшение мембранного потенциала) на мембране мотонейрона, с которым были соединены эти волокна. Экклс назвал такой градуальный потенциал возбуждающим постсинаптическим потенциалом (ВПСП). Как показано на рисунке в п. «Результаты» (слева), ВПСП снижают мембранный потенциал (деполяризация) до порогового уровня, повышая вероятность формирования потенциала действия.
Затем Экклс провел стимуляцию других сенсорных волокон — они изображены на рис. в п. «Процедура» (справа). Он создал гиперполяризующий градуальный потенциал (увеличение мембранного потенциала) на мембране принимающего мотонейрона. Экклс назвал такой градуальный потенциал тормозным постсинаптическим потенциалом (ТПСП). Как показано на рисунке в п. «Результаты» (справа), ТПСП увеличивают мембранный потенциал, препятствуя формированию потенциала действия.
ВПСП и ТПСП длятся лишь несколько миллисекунд, затем затухают, после чего происходит восстановление потенциала покоя. ВПСП связаны с активацией натриевых каналов, обеспечивающих приток ионов натрия. ТПСП возникают благодаря активации калиевых каналов, обеспечивающих отток ионов натрия (или благодаря открытию хлорных каналов и притоку ионов хлора).
Несмотря на то что величина градуального потенциала пропорциональна интенсивности стимула, на мембране тела мотонейрона не возникает потенциал действия, даже при значительной величине ВПСП. Причина проста: у большинства нейронов на мембране тела клетки отсутствуют потенциалзависимые ионные каналы. Стимул должен достигнуть начального сегмента — области, богатой потенциалзависимыми ионными каналами. Это лежащая поблизости от аксонного холмика или на аксонном холмике область, где возникает потенциал действия (Bender & Trussel, 2012).
а) Суммация. Мириады дендритных шипиков мотонейрона вносят свой вклад в мембранный потенциал, генерируя ВПСП и ТПСП. Как такие градуальные потенциалы взаимодействуют друг с другом на мембране? Что будет, если на мембране возникнет два ВПСП подряд? Важна ли длительность интервала между ними? Что происходит, когда ВПСП и ТПСП поступают одновременно?
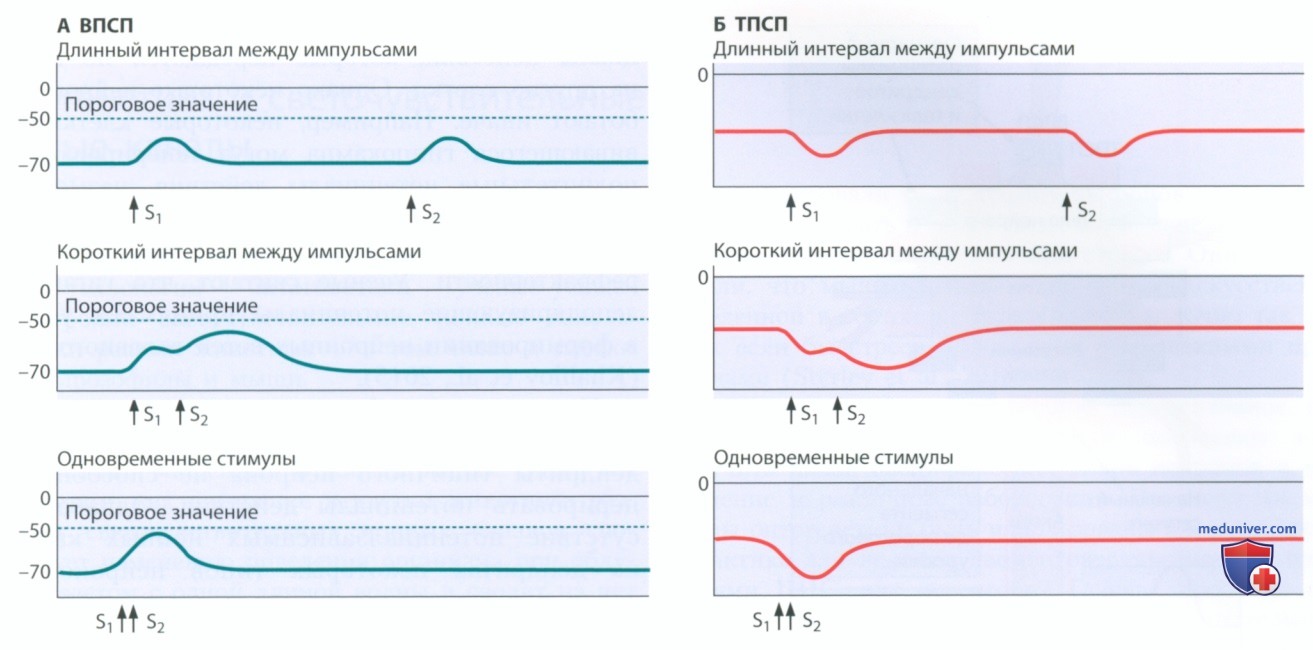
1. Временная суммация. Если за одним возбуждающим импульсом через некоторое время следует второй, регистрируется один ВПСП, а затем с задержкой регистрируется второй такой же ВПСП (рис. 1, слева вверху). Два разделенных длинным интервалом ВПСП являются независимыми и не взаимодействуют друг с другом. Если интервал между двумя ВПСП настолько мал, что они поступают непосредственно друг за другом, возникает один большой ВПСП (см. рис. 1, слева в центре).
P.S. Обычно нейроны одновременно принимают как возбуждающие, так и тормозные сигналы, суммируя их.
Здесь два возбуждающих импульса, поступающих в одну и ту же область, суммируются, обеспечивая большую деполяризацию мембраны, чем любой из них по отдельности. Взаимодействие между двумя ВПСП, поступающими с небольшой задержкой или одновременно (рис. 1, слева внизу), называют временной суммацией. В правой части рисунка представлены аналогичные схемы временной суммации для ТПСП. Временная суммация применима как к ВПСП, так и к ТПСП.
2. Пространственная суммация. Как пространственное разделение влияет на сигналы, поступающие на клеточную мембрану? Воспользовавшись двумя регистрирующими электродами (R1 и R2), мы можем узнать, как пространственные взаимоотношения влияют на суммацию сигналов. Если два ВПСП одновременно регистрируются на удаленных участках мембраны (рис. 2), они никак не влияют друг на друга. Если два ВПСП, возникающие практически одновременно, поступают на близлежащие участки мембраны, они суммируются, формируя ВПСП большей величины (рис. 2). Такая пространственная суммация возникает, когда два сигнала не разделены ни во времени, ни в пространстве. Аналогичным образом два одновременно возникающих ТПСП суммируются, если они не разделены ни в пространстве, ни во времени и не суммируются, если они поступают на расположенные далеко друг от друга участки мембраны.
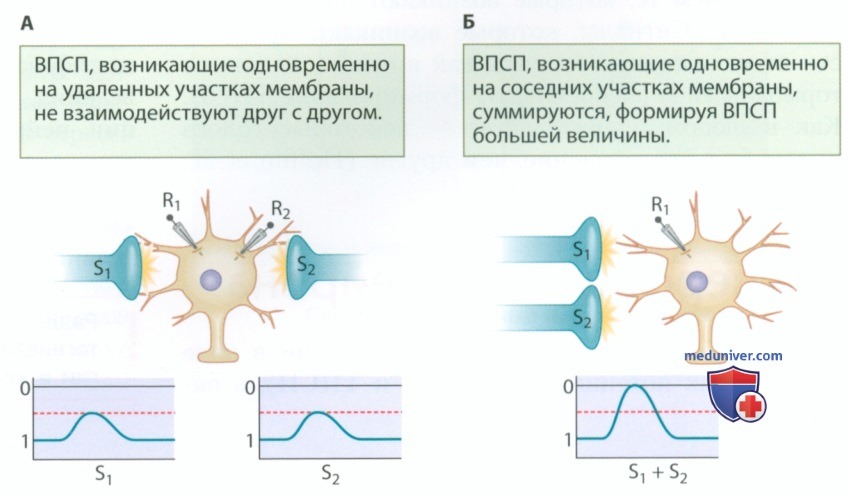
3. Роль ионов в суммации. Суммация применима к любым комбинациям ВПСП и ТПСП. Это имеет определенный смысл, если считать, что приток и отток ионов суммируются. Приток ионов натрия, сопровождающий формирование ВПСП, суммируется с притоком ионов натрия, обеспечивающим формирование второго ВПСП, если они не разделены во времени и пространстве. Если потоки ионов возникают в разное время на разных участках мембраны, суммация невозможна.
То же самое можно сказать о потоках ионов калия. Если они не разделены во времени и пространстве, то они суммируются. Если потоки ионов возникают в разное время на удаленных друг от друга участках мембраны, суммация отсутствует. Механизмы суммации ВПСП и ТПСП идентичны. Приток ионов натрия, сопровождающий формирование ВПСП, суммируется с оттоком ионов натрия, обеспечивающим формирование ТПСП, создавая разностный сигнал только в том случае, если входящие сигналы не разделены во времени и в пространстве. Если сигналы разделены во времени и пространстве, они не взаимодействуют друг с другом, поэтому суммация не происходит.
Ответ нейрона, принимающего тысячи сигналов от нестрональных клеток, ничем не отличается от ответа нейрона, принимающего лишь несколько сигналов — все нейроны суммируют не разделенные во времени и пространстве сигналы. Таким образом, мембрана тела нейрона всегда испытывает суммарное воздействие множества распределенных во времени или пространстве сигналов. Можно сказать, что нейрон анализирует поступающие извне сигналы, прежде чем дать ответ на их принятие. Окончательное решение принимает начальный сегмент аксона, на котором происходит формирование потенциала действия.
в) Потенциалзависимые ионные каналы и потенциал действия. В отличие от мембраны тела нейрона, мембрана аксона богата потенциалзависимыми ионными каналами, которые в том числе располагаются в начальном сегменте (рис. 3). Ионные каналы, как и каналы аксона кальмара, реагируют на определенное значение мембранного потенциала. Фактическая величина порогового мембранного потенциала зависит от типа нейрона, но мы, чтобы не усложнять, будем считать ее равной -50 мВ.
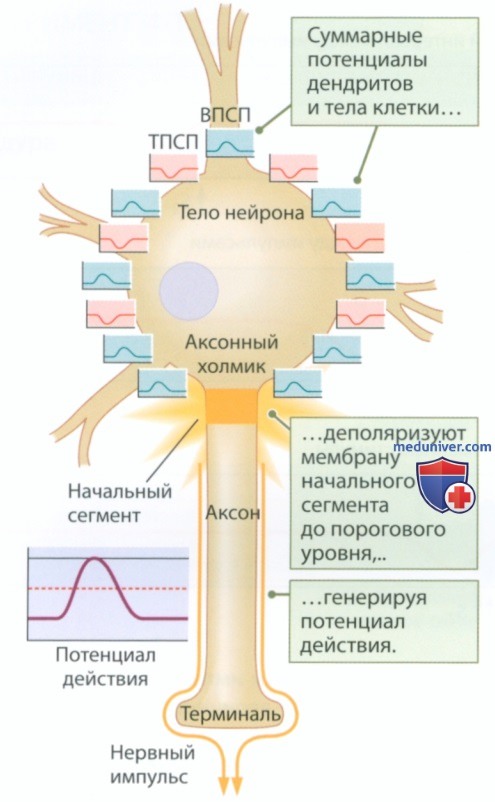
Чтобы создать потенциал действия, суммированные градуальные потенциалы (ТПСП и ВПСП) должны деполяризовать мембрану начального сегмента до -50 мВ. При краткосрочном отклонении мембранного потенциала до порогового значения потенциалзависимые ионные каналы открываются, обеспечивая формирование одного или нескольких потенциалов действия. Если пороговая величина мембранного потенциала сохраняется в течение более длительного периода, потенциалы следуют один за другим с частотой, которую определяет работа ворот потенциалзависимых каналов. Затем каждый потенциал действия распространяется, формируя нервный импульс, который проходит от начального сегмента по всей длине аксона.
У многих нейронов сильно ветвящиеся дендриты, однако на дендритах мало потенциалзависимых ионных каналов, поэтому обычно они не генерируют потенциалы действия. Расположенные на большом расстоянии друг от друга ветви дендритов могут в меньшей степени влиять на генерируемые начальным сегментом потенциалы действия, чем расположенные на более близком расстоянии ветви. Соответственно, сигналы, возникающие ближе к начальному сегменту, обычно имеют большее влияние, чем те, которые возникают на некотором отдалении. Сигналы, которые возникают поблизости от начального сегмента, как правило, являются тормозными и обеспечивают формирование ТПСП. Как в любом правительстве — некоторые голоса имеют большее значение, чем другие (Hofflin et al., 2017).
г) Универсальность нейрона. Дендриты осуществляют сбор информации в виде градуальных потенциалов (ВПСП и ТПСП), а начальный сегмент генерирует дискретные потенциалы действия, которые передаются по аксону на другие клетки. Однако некоторые нейроны работают иначе. Например, некоторые клетки развивающегося гиппокампа могут генерировать дополнительные потенциалы действия, называемые гигантскими деполяризующими потенциалами, в период, когда клетка обычно находится в состоянии рефрактерности. Ученые считают, что гигантские деполяризующие потенциалы принимают участие в формировании нейронных сетей головного мозга (Khalilov et al., 2015).
Поскольку мембрана тела нейрона не содержит потенциалзависимых ионных каналов, дендриты типичного нейрона не способны генерировать потенциалы действия. Однако присутствие потенциалзависимых ионных каналов на дендритах некоторых типов нейронов позволяет им генерировать потенциалы действия. Распространение потенциала действия от начального сегмента к дендритам нейрона называют обратным распространением. Обратное распространение, которое информирует дендриты о том, что нейрон распространяет потенциал действия по аксону, может играть некую роль в пластических изменениях нейронов, лежащих в основе обучения. Например, обратное распространение может стать причиной невосприимчивости дендритов к поступающим сигналам, сделать дендриты электрически нейтральными или обеспечить усиление поступающих на отдельные дендриты сигналов (Schiess et al., 2016).
P.S. Принцип 2. Нейропластичность — отличительный признак функционирования нервной системы.
Нейроны некоторых не относящихся к млекопитающим видов не имеют дендритов. А некоторые ионные каналы реагируют не на изменение мембранного потенциала, а на свет. Многочисленные различия между нейронами показывают, что нервная система использует встречающиеся у разных видов животных структурные и функциональные модификации для производства адаптивного поведения. Чтобы изучить специфические функции нейрона, нейробиологи встроили в нейроны определенных типов светочувствительные ионные каналы — об этом можно прочитать ниже.
д) Исследование. Оптогенетика и светочувствительные ионные каналы. Способные реагировать на свет ионные каналы были обнаружены у некоторых не относящихся к классу млекопитающих видов животных. С помощью трансгенной технологии — оптогенетики — ученые успешно встроили светочувствительные ионные каналы в клетки разных животных — в том числе в клетки червя, дрозофилы и мыши.
Оптогенетика предполагает применение генетических методов и воздействия светом для управления клетками-мишенями в живых тканях. Здесь мы рассмотрим, как внедрение в клетки животного различных светочувствительных ионных каналов способствует изменению поведения организма при облучении светом с одной длиной волны и сводит на нет подобные изменения при облучении светом с другой длиной волны.
Примером светочувствительного ионного канала является полученный из зеленой водоросли Chlamydomonas reinhardtii ченнелродопсин-2 (ChR2). Светочувствительный ионный канал ChR2 реагирует на синий свет, ненадолго открываясь и пропуская ионы Na+ и К+. Возникающая деполяризация возбуждает клетку, заставляя ее генерировать потенциалы действия.
Галородопсин (NpHR) — это светочувствительный ионный насос, который отвечает за транспорт ионов хлора. Он содержится в клетках архей, называемых галобактериями. При облучении зелено-желтым светом NpHR закачивает ионы хлора в клетку, обеспечивая гиперполяризацию и подавляя активность клетки.

Поведение животных, в мембраны клеток которых встроены светочувствительные ионные каналы, можно контролировать, подвергая клетки их нервной системы воздействию света с определенной длиной волны. Методы оптогенетики позволяют встроить светочувствительные ионные каналы в специфические нейронные сети таким образом, чтобы свет стимулировал только определенную группу нейронов.
Стресс влияет как на поведение, так и на гормональный статус организма, передаваясь другим членам социума. Воспользовавшись оптогенетическими методами, Стирли и его коллеги (Sterley, 2018) сумели подавить активность специфической группы нейронов гипоталамуса мыши при стрессе, таким образом предупредив обычно возникающие из-за стресса изменения в мозге. Затем они сделали следующий шаг — подавили активность нейронов партнера находящейся в состоянии стресса мыши, благодаря чему стресс не передался ему. После этого группа исследователей провела обратный эксперимент: в отсутствие стрессовых факторов они оптогенетическими методами активировали ту же группу нейронов гипоталамуса, чтобы вызвать в мозге такие же изменения, как те, которые обычно возникают при стрессе. Они обнаружили, что мышь-партнер ведет себя с искусственно введенной в состояние стресса мышью точно так же, как если бы стресс был вызван естественными причинами (Sterley et al., 2018).
Оптогенетика — чрезвычайно перспективное направление; методы оптогенетики позволяют нам изучить вклад специфических типов нейронов в поведение и различные заболевания. Но могут ли методы оптогенетики быть использованы в клинической практике для лечения пациентов с такими заболеваниями ЦНС, как депрессия? Главная проблема состоит в том, что для переноса светочувствительных ионных каналов в нейроны определенных областей мозга используют специфические вирусы, а применение вирусов в человеческой популяции все еще связано с определенными сложностями. Результаты ряда исследований говорят о том, что связанные с утратой светочувствительных клеток сетчатки нарушения зрения можно скорректировать, внедрив светочувствительные ионные каналы в выжившие нейроны сетчатки (Pan et al., 2015).
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) - профессор, д.м.н. П.Е. Умрюхин
- Читать далее "Доставка информации в нервную систему и обратно"
Редактор: Искандер Милевски. Дата публикации: 12.7.2023
- История поиска источников электрической активности в нервной системе
- Мембранный потенциал: потенциал покоя, потенциал действия, нервный импульс, рефрактерный период
- Как нейроны интегрируют информацию: суммация, универсальность нейрона
- Доставка информации в нервную систему и обратно
- Резюме по использованию электрических сигналов нейронами для передачи информации