Начальные стадии пикорнавирусных инфекций. Поражения пикорнавирусами
Пикорнавирусы человека размножаются только в клетках человека и обезьяны, так как лишь эти клетки несут специфические рецепторы, на которых адсорбируется вирус (обзоры Холленда и Хойер, 1962; Холленда, 1964). Рецепторы были выделены из плазматических мембран чувствительных клеток, но очистить их пока не удалось. Показано, что они содержат белок, прочно связанный с мембранными липидами.
Клетки амниона человека, невосприимчивые к полиовирусной инфекции in vivo, до эксплантации in vitro не имеют специфических рецепторов, но эти рецепторы появляются после периода культивирования (Холленд, 1961; Шани и др., 1966). Были также получены данные (М. Тейлор и др., 1971) о том, что энтеровирус крупного рогатого скота избирательно разрушает определенные злокачественные опухоли мышей in vivo, что объясняется наличием в раковых клетках рецепторов, которые не проявляются в других мышиных тканях.
Рецепторы для различных энтеровирусов, по-видимому, отличаются друг от друга, что подтверждается опытами по перекрестному насыщению, различной чувствительностью к блокированию антиклеточной сывороткой и к протеазам, а также различным временем регенерации при 37°С (см. Левит и Кроуэл, 1967).
«Нечувствительные» клетки млекопитающих можно заразить энтеровирусной нуклеиновой кислотой; эффективность бляшкообразования в этом случае чрезвычайно низка, однако вирусное потомство содержит совершенно нормальные инфекционные вирионы (Холленд, 1964). Для введения полиовируса в клетки, лишенные специфических рецепторов, используют слияние клеток под действием инактивированного вируса Сендай; вирус в таких клетках размножается нормально (Эндерс и др., 1971).
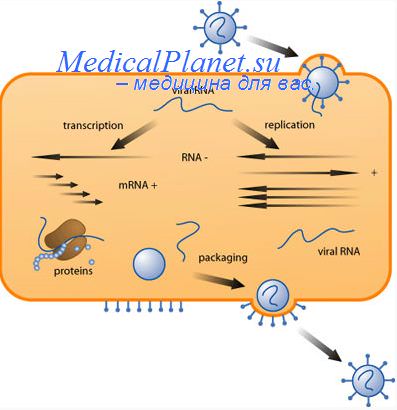
Раздевание пикорнавирусов начинается, когда адсорбированный вирион еще находится вне клетки. Йоклик и Дарнелл (1961) показали, что около половины полиовирионов, прикрепившихся к клеткам при 4°С, при повышении температуры до 37°С элюируется в среду; элюировавшиеся вирионы претерпевали такие изменения капсида, которые делали их неустойчивыми к высоким солевым концентрациям и неспособными к реадсорбции на чувствительных клетках.
Мэндел (1971а) изучил влияние этого зависимого от температуры процесса на конформационное состояние капсида; оказывается, физические свойства вирионов сильно изменяются, так что они не могут эффективно связывать нейтрализующие антитела. Кроуэлл и Филлипсон (1971) показали, что элюировавшиеся вирионы Коксаки ВЗ, неинфекционные из-за неспособности прикрепляться к клеткам HeLa, избирательно теряли полипептид VP4; они предположили, что VP4 —первый белок, который высвобождается в процессе раздевания.
Этого может быть достаточно для того, чтобы ослабить структуру капсида и тем самым обеспечить его автоматический распад при вхождении в цитоплазму.
Холл и Рюккерт (1971) показали, что в случае кардиовирусов процесс внеклеточного раздевания идет еще глубже, При 37°С более половины адсорбированных вирионов мышиного энцефаломиелита освобождает свою РНК в среду; капсиды временно остаются присоединенными к плазматической мембране, а затем диссоциируют на 145-пентамеры.
Внеклеточное раздевание происходит только при 37 °С и подавляется в щелочных (рН 8,5) или гипертонических (40 мМ MgCl2) растворах, т. е. при условиях, которые, как было ранее показано, стабилизируют вирионы мышиного энцефаломиелита (ME) против тепловой инактивации, способствуют усилению фагоцитоза (Дейлс, 1965а) и увеличивают эффективность бляшкообразования при заражении клеток изолированной вирусной РНК (обзор Пагано, 1970). Освобождение РНК в среду само по себе, очевидно, не имеет прямого отношения к естественному заражению, но разрушение капсида, делающее возможным освобождение РНК, по-видимому, является первым этапом раздевания вириона.
Не совсем ясно, каким именно образом вирион проходит через плазматическую мембрану. Дейлс и др. ( 1965а) представили доказательства цитоза, однако Даннебэк и др. (1969) предполагают, что происходит прямое проникновение адсорбированного вируса в цитоплазму. Неизвестно также, когда происходит удаление VP4 и освобождение РНК: перед тем ли, как вирион войдет в цитоплазму (Чэн и Блэк, 1970), или после связывания с мембраной фагоцитарной вакуоли (Мэндел, 1967).
- Читать далее "Трансляция пикорнавирусов. Этапы трансляции пикорнавирусов"
Оглавление темы "РНК вирусы. Пикорнавирусы":1. Размножение РНК вирусов. Транскрипция и трансляция РНК вирусов
2. Репликация РНК вирусов. Почкование РНК вирусов
3. Регуляция РНК вирусов. Активность РНК вирусов
4. Синтез РНК вирусов. Цикл размножения пикорнавирусов
5. Начальные стадии пикорнавирусных инфекций. Поражения пикорнавирусами
6. Трансляция пикорнавирусов. Этапы трансляции пикорнавирусов
7. Синтез молекул РНК пикорнавирусов. Этапы синтеза РНК пикорнавирусов
8. Регуляция синтеза РНК пикорнавирусов. Ферменты синтеза РНК пикорнавирусов
9. Сборка пикорнавирусов. Этапы сборки пикорнавирусов
10. Регуляция выражения генов пикорнавирусов. Механизмы представления генов пикорнавирусов