Рекомбинация вирусов ВИЧ
Во время начальных процессов взаимодействия вирусов с клетками существует возможность рекомбинации между геномами двух различных вирусов, инфицирующих одну и ту же клетку. Этот процесс можно зафиксировать, только если в одну клетку проникли нуклеоиды двух гетеротипических вирусов (т.е. вирусов, у которых различаются их геномные РНК).
При этом могут образовываться новые рекомбинантные вирусы, обладающие новыми биологическими и патогенными свойствами. В вновь выделенном изоляте ВИЧ факт его рекомбинантного происхождения определяют по появлению генетических различий в специфических областях вирусного генома (например, в Gag и Env).
Первый изолят ВИЧ-1, который идентифицировали как рекомбинантную форму, назвали MAL (Н2231). Этот вирус выделили из образца крови, полученной из Демократической Республики Конго и хранящейся с 1976 года; вирус представляет собой рекомбинант между ВИЧ-1 субтипов А и D (63). В настоящее время от 10 до 40% клинических изолятов, выделяемых в Африке, представлено циркулирующими рекомбинантными формами (CRFs), что говорит о достаточно широком распространении рекомбинантных вирусов (L. Demeter, персональное интервью).
По-видимому, рекомбинация может происходить в любой области вирусного генома, но особенно часто она затрагивает гены gag и env. Рекомбинацию регистрируют преимущественно между вирусами разных субтипов.
В изолятах ВИЧ-1 из Китая была зафиксирована рекомбинация между вирусами субтипов В и С в области гена tat и между субтипами В и F в различных областях генома. В настоящее время известно, что бывшие субтипы Е и I представляют собой рекомбинантные формы.
В рекомбинантных вирусах из Южной Америки обнаружены признаки рекомбинации между субтипами A, G, J и К и другими неизвестными квазивидами. Очевидно, рекомбинацию можно распознать, только если области, в которых происходят точечные разрывы, не нарушают экспрессии генов.
Недавние исследования показали, что рекомбинация, происходящая in vitro между вирусами в пределах одного субтипа, например между вирусами субтипа С или вирусами субтипа В, происходит с одинаковой частотой, но частота рекомбинации между вирусами различных субтипов, например между В и С гораздо ниже.
По-видимому, такое уменьшение частоты случаев рекомбинации между вирусами разных субтипов связано с различиями в трехнуклеотидной последовательности в сигнальной области инцициации димеризации у вирусов субтипов В и С. Рекомбинация, очевидно, зависит от согласованных последовательностей в сигнальной области инцициации димеризации. Тем не менее были обнаружены даже вирусы, образовавшиеся путем рекомбинации вирусов различных групп ВИЧ-1 (М, N и О).
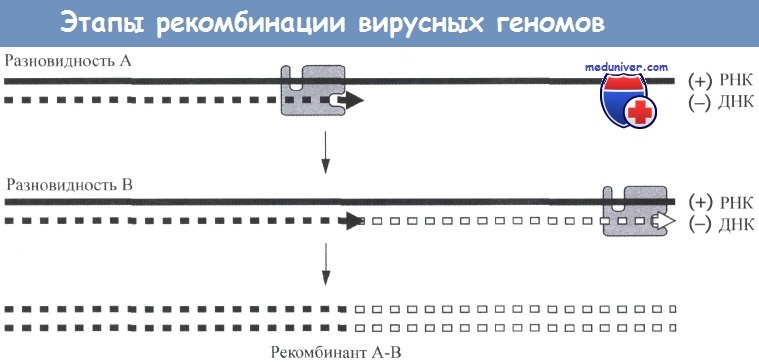
Данная схема иллюстрирует процесс образования рекомбинантов во время обратной транскрипции путем переноса цепей нуклеиновых кислот.
Показаны плюс-нити геномных РНК двух генетически различных разновидностей ВИЧ, совместно инфицирующих клетку.
В приведенном на рисунке примере обратная транскриптаза (обозначенная в виде серого прямоугольника) инициирует синтез, используя в качестве матрицы плюс-цепь геномной РНК разновидности А.
По мере синтеза могут возникать условия, вызывающие перенос обратной транскриптазы и синтезируемой минус-нити ДНК, копированной с разновидности А на геномную РНК разновидности В.
Затем продолжается синтез, при котором в качестве матрицы используется геномная РНК разновидности В (показана в виде пунктирной линии).
Образующаяся в результате провирусная ДНК содержит информацию в виде последовательностей нуклеотидов от обеих разновидностей вирусов - А и В.
Представленный на рисунке процесс носит название переноса цепи и может осуществляться обратной транскриптазой in vitro.
Было идентифицировано несколько рекомбинантных вирусов, образованных из трех различных субтипов. Некоторые из таких вариантов ВИЧ-1 образуются в результате повторных рекомбинаций (например, рекомбинация рекомбинантной формы CRF07-BC с другими штаммами). Однако рекомбинация между ВИЧ-1 и ВИЧ-2, по-видимому, является невозможной, поскольку шпилеобразные участки инициации образования димеров их РНК существенно различаются между собой и не могут образовывать гетеродимеры РНК, как необходимое условие рекомбинации.
Как отмечали выше, рекомбинация может происходить при одновременном инфицировании клетки более чем одним вирусом или при суперинфицировании клетки, которое успевает произойти до прекращения экспрессии CD4 в результате первичной инфекции и наступления резистентности к последующему инфицированию. Этот феномен может происходить на ранних стадиях инфекционного процесса (в первые 1-3 дня), в латентно инфицированной клетке, в клетке, содержащей дефектный вирус, или в клетках, инфицированных вирусами, которые не вызывают снижения экспрессии CD4-белка на поверхности клетки.
Некоторые исследования показали, что в случае ВИЧ-1 акты рекомбинации могут происходить два - три раза на геном на один цикл репликации, т.е. частота рекомбинаций ВИЧ-1 гораздо выше, чем у других ретровирусов. С помощью метода, в котором используют различные т.н. репортерные вирусы in vitro, было показано, что в Т-лимфоцитах на один цикл репликации ВИЧ в среднем приходится девять рекомбинаций на один вирусный геном, а в инфицированных макрофагах число рекомбинаций еще больше. В других исследованиях выявленная частота генетических рекомбинаций ВИЧ-1 в Т-клетках и в макрофагах примерно одинакова.
Существенно, что в в процессе эффективной деградации РНК-матрицы под действием фермента РНКазы Н (что, вероятно, происходит во время транскрипционной паузы) возникают молекулярные условия, благоприятные для рекомбинации. Следовые количества нуклеотидных гомополимеров вызывают короткую остановку в работе фермента обратной транскриптазы, и в этот момент может происходить переключение ДНК-полимеразы на другую матрицу РНК, что и приводит к созданию рекомбинантных форм геномов вирусов.
Некоторые исследования in vitro показали, что рекомбинация в области гена Gag происходит во время обратной транскрипции в начальном участке кодирующей области, необычно обогащенной по гуанину (G), и вызывает паузы в работе обратной транскриптазы. Другие исследования позволили предположить наличие локальных «горячих точек» рекомбинации на протяжении генома ВИЧ.
По-видимому, такой «горячей точкой» является последовательность нуклеотидов, кодирующая С2-домен оболочечного белка-предшественника. Кроме того, вторичная структура РНК (а именно, шпилечные структуры) может быть особенно благоприятна для рекомбинаций. К тому же мутации замены G на А происходят с высокой частотой в участках нуклеиновых кислот с высоким содержанием остатков G. В обогащенных по G областях происходят с повышенной частотой как рекомбинации, так и мутации с заменой G на А (что, возможно, связано с активностью APOBEC3G).
Процесс гомологичной рекомбинации может привести к тому, что два дефектных вируса породят рекомбинантную форму с цитопатогенными свойствами/ Рекомбинации становятся причиной появления новых штаммов в популяции, таким образом осложняя лечение (из-за устойчивости к лекарственным препаратам) и разработку вакцин. В одном эксперименте с инокуляцией обезьяне двух мутантных штаммов SIV, обладающих низкой репликативной способностью, через 2 недели был выделен быстрореплицирующийся рекомбинантный вирус.
В другом исследовании отобранные мутантные вирусы SIV, несущие делеции в генах vpx, vpr и nef, вводили в организм одновременно или последовательно путем внутривлагалищной инокуляции. Рекомбинации происходили in vivo . Однако если первым был введен мутан-тный вирус, обладающий высокой скоростью репликации, образования рекомбинантных форм не обнаружили. Последний факт, скорее всего, связан с доминированием первого вируса и отсутствием суперинфицирования.
- Читать "Основные моменты острого заражения ВИЧ"
Оглавление темы "Острое инфицирование ВИЧ":- Скорость репликации ВИЧ - кинетика
- Влияние гликолизирования белков стенки ВИЧ на выбор инфицируемых клеток
- Экспрессия CD4 белка клетками после инфицирования ВИЧ
- Изменения клеток in vitro после инфицирования ВИЧ
- Суперинфекция ВИЧ на клеточном уровне
- Множественная ВИЧ-инфекция - заражение разными вирусами
- Частота (распространенность) суперинфекции ВИЧ
- Факторы влияющие на суперинфицирование ВИЧ
- Рекомбинация вирусов ВИЧ
- Основные моменты острого заражения ВИЧ