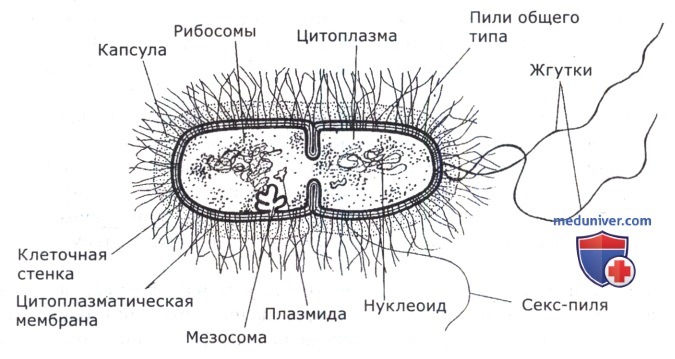
Способы адаптации бактерий к условиям существования
Возможность микроорганизмов к существованию в определенных условиях выработана в процессе эволюции. Выше перечислены физические, химические и биологические факторы, которые оказывают влияние на жизнедеятельность микроорганизмов. Способы адаптации к некоторым из них заключаются в капсулои слизевыделении, спорообразовании, формировании покоящихся «некультивируемых» форм, трансформации в L-формы, адгезии к субстрату, в детоксикации, синтезе запасных питательных веществ и др. Рассмотрим их подробнее.
а) Капсулы и слизь. Капсулы (макро- и микрокапсулы) состоят из полисахаридов (например, у пневмококка) или полипептидов (например, у бацилл сибирской язвы). Они не являются обязательными компонентами клеток и образуются патогенными бактериями при попадании в макроорганизм как средство защиты от фагоцитоза. Слизистый слой, располагающийся поверх клеточной стенки, образуется в естественных условиях при формировании клеточных сообществ («мат», «слизистый тяж»), что придает им большую резистентность к неблагоприятным воздействиям.
б) Спорообразование. Споры —покоящиеся формы жизненного цикла грамположительных палочковидных бактерий. Они образуются внутри цитоплазмы (эндоспоры) при различных неблагоприятных условиях внешней среды и способны очень длительное время сохранять жизнеспособность (роды Bacillus, Clostridium, Sporosarcina, Desulfotomaculum и др.). Споры устойчивы к нагреванию, радиации, давлению, химическим веществам.
В них происходит репрессия генома, прекращаются метаболические процессы, уменьшается количество свободной воды, образуется дипиколиновая кислота и повышается концентрация ионов кальция. Процесс споруляции начинается в конце стационарной фазы роста с удвоения нуклеоида, который отходит к одному из полюсов клетки. Затем происходит инвагинация цитоплазматической оболочки, и клетка разделяется на два неравных протопласта, каждый из которых содержит нуклеоид. Меньший из протопластов (проспора) покрывается второй оболочкой, являющейся ЦПМ материнской клетки.
Между двумя листками мембраны споры формируется основной слой — кортекс, состоящий из пептидогликана, имеющего сетчатую структуру. Снаружи зрелая спора покрывается толстой, иногда многослойной оболочкой. Поверх этой оболочки у некоторых бактерий образуется еще один рыхлый слой — экзоспориум. Процесс образования споры продолжается 18-20 ч.
При попадании споры в благоприятные условия происходит ее активация, заключающаяся в дерепрессии генома и мобилизации метаболических процессов с последующим прорастанием в вегетативные клетки. Спора активно поглощает воду, набухает, у нее усиливается дыхание и возрастает активность ферментов, выделяются аминокислоты, дипиколиновая кислота, пептиды, разрушается кортекс и теряется термоустойчивость. Оболочка споры разрывается, и из нее выходит ростовая трубка. Прорастание споры происходит быстрее, чем ее образование, и занимает 4-5 ч.
У палочковидных бактерий образуется по одной споре, которая может располагаться центрально (В. anthracis), субтерминально (С. botulinum) или терминально (С. tetani). Спора имеет овальную или круглую форму, ее диаметр может превышать диаметр самой бактерии (клостридии).
в) Покоящиеся (некультивируемые) формы образуются при остром недостатке питательных веществ. При этом клетка уменьшается, принимает округлую форму. Возможно образование «фильтрующих» форм, способных проходить через бактериальный фильтр. Количество РНК и рибосом уменьшается, снижается интенсивность метаболизма. В клеточной стенке появляются белки «голодания», что сопровождается изменением антигенной структуры, снижается вирулентность. Способность к росту на питательных средах резко уменьшается, вплоть до временного полного его прекращения. В таком состоянии бактерии длительно сохраняют жизнеспособность.
В благоприятных условиях происходит реверсия к вегетативному состоянию. Считают, что процесс перехода бактерий в состояние покоя находится под контролем особого гена. Некультивируемые формы обнаружены у бактерий родов Escherichia, Salmonella, Shigella, Legionella, Listeria и др.
г) L-формы. Трансформация в L-формы обусловлена частичной или полной утратой способности к синтезу пептидогликана. При этом бактерии сохраняют способность к длительному переживанию и размножению в макроорганизме и на питательных средах. Их размеры уменьшаются, и появляется способность проходить через бактериальные фильтры. Биохимическая активность, вирулентность, иммуногенность у них ниже, а устойчивость к антителам, антибиотикам, лизоциму выше, чем у аналогичных исходных культур.
L-формы впервые получены в Институте Листера в 1935 г. у Streptobacillus moniliformis. Позже было установлено, что пенициллин и его производные, а также ристоцетин, ванкомицин, новобиоцин, бацитрацин, некоторые аминокислоты и др. нарушают процесс образования клеточной стенки у микроорганизмов. Происходит потеря ряда поверхностных антигенов, теряется чувствительность не только ко многим антибиотикам (к эритромицину чувствительность сохраняется), но и к антителам, бактериофагам и бактериоцинам.
Различают стабильные и нестабильные L-формы. Первые легко реверсируют в исходные бактериальные формы после удаления трансформирующего агента и создания оптимальных условий культивирования. Поэтому образование L-форм считается одним из проявлений адаптации микроорганизмов к неблагоприятным условиям существования. Стабильные формы утрачивают способность к реверсии. L-формы часто обнаруживают при хронических инфекциях (бруцеллез, ревматизм и др.), бактерионосительстве.
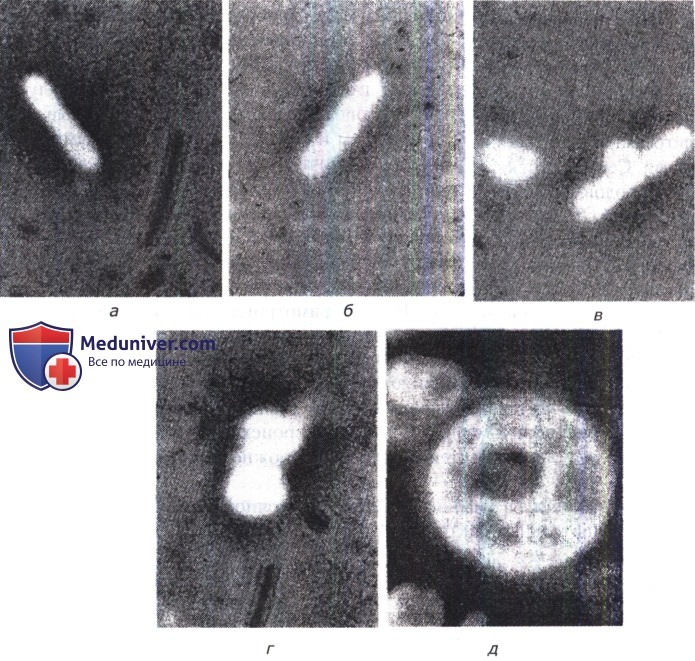
а —клетка Proteus sp. после 30 мин контакта с пенициллином;
б — та же клетка после 60 мин контакта с пенициллином (клеточная стенка теряет упругость, становится мягкой, и цитоплазма внедряется в нее в центре клетки);
в — после 90 мин контакта с пенициллином (цитоплазма, которая переместилась под ЦПМ, приобретает округлую форму);
г — после 120 мин контакта с пенициллином форма клетки изменилась, образовались сферопласты, которые размножились — зарегистрирована фаза L-роста Proteus sp.;
д — через 5 ч контакта с пенициллином в гипотонической среде: один из сферопластов набух, формируя гигантскую клетку с большими вакуолями.
д) Таксис — направленное движение подвижных бактерий по градиенту концентраций эффекторов (аттрактантов) или против них. Такое направленное движение помогает бактериям выбирать источники питания и наиболее благоприятные экологические ниши, избегать высокой концентрации реппелентов (веществ, вызывающих отрицательный таксис). Формы таксиса зависят от раздражителя: хемотаксис, фототаксис, аэротаксис, термотаксис и др. Так, хемоаттрактантами для кишечной палочки являются аминокислоты, моносахара, реппелентами — спирты, кислоты, сероводород, индол и др.
Таксис достигается при помощи специфических рецепторов — метилакцептирующих белков (МАБ), принимающих или подавляющих сигнал таксиса, который передается базальным тельцам жгутиков. Способность к таксису генетически детерминирована. Специальные гены кодируют белки, образующие локомоторный аппарат, белки и ферменты, участвующие в преобразовании сигналов.
е) Адгезия — способность прикрепляться, прилипать к субстрату; она обеспечивается наличием у микроорганизмов пилей общего типа, покрывающих поверхность клеток белков, наружной мембраны, ЛПС и др. Адгезия является начальной фазой любого инфекционного процесса. Она позволяет возбудителям приживаться в организме, вызывать колонизацию клеточных поверхностей.
Адгезины помогают бактериям образовывать биопленки, состоящие из популяций разных видов микроорганизмов, удерживать их вместе. Биопленки выполняют функцию защиты и питательного косубстрата в естественной среде обитания. Образование адгезинов находится под контролем плазмид.
ж) Запасные питательные вещества — это продукты клеточного метаболизма, образующиеся при избытке экзогенных питательных веществ в условиях заторможенного роста. Они используются при голодании или в период адаптации микроорганизмов к новым субстратам. Запасные вещества присутствуют в клетке в осмотически инертной форме, они нерастворимы в воде, но способны включаться в метаболизм. Основной их функцией считают обеспечение бактериальных клеток энергией и необходимыми элементами — углеродом, фосфором. Главными запасными веществами являются полисахариды (крахмал, гликоген, гранулеза), липиды (поли-р-оксибутират — «суданофильные» гранулы), полифосфаты (зерна волютина), сера.
Они присутствуют в клетках в виде сферических или неправильной формы гранул, окруженных оболочкой. Запасание полисахаридов присуще бактериям кишечной группы, стрептококкам, клостридиям, дрожжам и др. Запасные липиды обнаружены у бактерий родов Bacillus, Azotobacter, Micrococcus. Накопление волютина широко распространено у бактерий родов Corynebacterium, Micrococcus, Lactobacillus и др.
Детоксикационная способность микроорганизмов позволяет им выживать в присутствии некоторых токсических веществ (например, солей тяжелых металлов), антибиотиков, дезинфектантов и др. Механизм такого действия связан с восстановлением веществ до нетоксичных соединений, ферментативным разрушением, изменением проницаемости клеточных стенок, уменьшением чувствительности молекул-мишеней к токсическим веществам или энергозависимому выбросу ксенобиотиков из клетки.
- Читать далее "Пигменты микроорганизмов и фотосинтез"
Редактор: Искандер Милевски. Дата публикации: 18.05.2019