L-формы бактерий (L-трансформация)
Явление L-трансформации бактерий впервые было описано E.Klieneberger в 1935 г. Свое название L эти формы получили в честь Листеровского института в Лондоне, где они были открыты. С тех пор накопилось большое количество данных, свидетельствующих о том, что L-форма — это одна из форм существования бактерий в природе.
В нашей стране проблема L-трансформации бактерий изучалась В.Д. Тимаковым, ЕЯ. Каган, С.В. Прозоровским и их многочисленными учениками и последователями. По мнению исследователей, эта проблема имеет два аспекта: общебиологический и медицинский. Общебиологический аспект определяется принципиальной возможностью существования бактерий в виде обычных бактериальных форм с клеточной стенкой и L-форм, лишенных ее.
Медицинский аспект определяется ролью L- форм в инфекционном процессе.
Феномен L-трансформации наблюдается у многих бактерий, касается различных признаков и свойств и у разных видов бактерий довольно однотипен. По результатам гибридизации нуклеиновых кислот и по нуклеиновому составу выявляется родство L-форм с исходными бактериями.
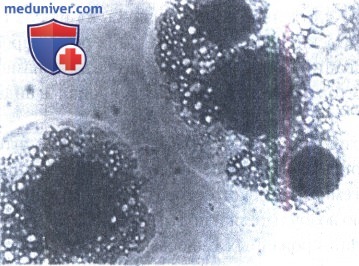
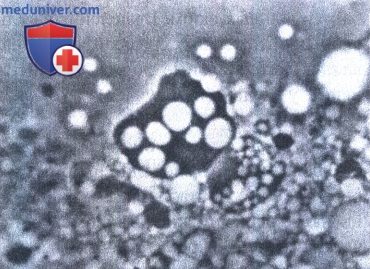
L-формы бактерий, частично или полностью лишены клеточной стенки и имеют только цитоплазматическую мембрану. Колонии L-форм разных видов бактерий сходны по морфологии. Некоторые вариации зависят от фазы роста, условий и среды культивирования, но не от видовых особенностей, Как правило, колонии имеют нежный, ажурный, сильно вакуолизированный край и врастающий в агар центр. Они хорошо видны невооруженным глазом. Микроструктурные элементы колоний разнообразны:
1) сферические тела, варьирующие по форме, размеру и оптической плотности;
2) элементарные тельца или гранулы, лежащие свободно или внутри сферических тел и вакуолей;
3) аморфные бесструктурные массы различной конфигурации;
4) извитые и нитевидные структуры.
L-формы могут быть стабильными, т.е. способными сохраняться и перевиваться независимо от наличия в среде L-трансформирующих агентов, что связано с необратимой утратой определенных звеньев биосинтеза клеточной стенки и в первую очередь основных компонентов пен гидогликана — диаминопимелиновой и Мурановой кислот. У них нарушается синтез глюкозамина. Иногда рост стабильных L-форм зависит от трансформирующего агента (например, у пенициллинозависимых L-форм стрептококка). Нестабильные L-формы при культивировании на средах без L-трансформирующего фактора реверсируют в исходные бактерии.
Патогенность стабильных L-форм может быть обусловлена как сохранением способности продуцировать некоторые факторы патогенности, так и приобретением новых факторов патогенности. Патогенность нестабильных L-форм связана с их способностью реверсировать в исходные патогенные виды и индуцировать развитие хронических инфекционных процессов.
Способность стабильных L-форм вызывать инфекционный процесс показана на различных экспериментальных моделях (менингоэнцефалит кроликов, ангина обезьян, экспериментальный листериоз ягнят и кроликов).
L-формы удавалось выделять из крови и органов больных людей при инфекционных процессах, вызванных бактериальными формами. Во всех случаях отмечали длительную персистенцию возбудителя и, как правило, менее тяжелое, но хроническое течение заболевания.
В качестве примеров инфекционных процессов, вызываемых стабильными L-формами, можно привести следующие:
1. Туберкулезная инфекция. Было показано, что от больных туберкулезом L-формы выделяют в виде чистых культур при полном отсутствии классических бактериальных форм. Выделенные культуры длительно пассировались на искусственных питательных средах и в организме морских свинок, часть из них реверсировала в бактериальные формы. При исследовании остаточных туберкулезных изменений у людей (60 случаев) у 75% пациентов возбудитель персистировал в виде L-формы.
2. Стрептококковая инфекция. При септическом эндокардите и ревмокардите от больных часто выделяли L-формы стрептококка. При экспериментальном инфицировании обезьян стабильными L-формами гемолитического стрептококка группы А наблюдали ангину, длительную персистенцию возбудителя и подъем титров специфических антител в крови. У белых мышей, инфицированных L-формами стрептококка, отмечали прогрессирующие патологические изменения в миокарде и развитие гломерулонефрита. Антигены L-форм обнаруживали в сердце, почках и особенно часто в лимфоидной ткани в течение 1 года (срок наблюдения).
3. Брюшнотифозная инфекция. При экспериментальной брюшнотифозной инфекции кроликов через 6-18 месяцев после заражения от всех животных выделяли бактерии с дефектом клеточной стенки (измененные и увеличенные бактериальные клетки, нестабильные культуры L-форм), частота их выделения из разных органов была в 3-8 раз выше, чем типичных бактерий. Таким образом было показано, что в процессе брюшнотифозной инфекции и бактерионосительстве имеет место образования L-форм.
4. Другие инфекции. Для возбудителей ряда других инфекций (менингоккока, гонококка, сальмонелл, листерий, вибриона холеры) показана возможность существования в организме больных в виде L-форм. Однако, если в случае возбудителя туберкулеза и стрептококковой инфекции значение L-форм в их персистенции не вызывает сомнений, то в отношении других микроорганизмов этот вопрос остается нерешенным.
Так, в экспериментах на животных и при изучении материала от больных исследователи часто встречались со смешанными популяциями бактерий и их L-форм, что значительно усложняло выяснение собственной роли L-форм в персистенции и инфекционном процессе. Отсутствие универсальной селективной модели для активного размножения, накопления биомассы и трудности выделения L-форм от больных затрудняют решение вопроса об их роли в персистенции возбудителей. Существуют большие методические трудности в решении вопроса о роли L-форм в инфекционном процессе, вызванном различными возбудителями. Система доказательств является достаточно сложной и требует усовершенствования и унификации.
Для выделения, индукции и культивирования L-форм необходимо правильно подобрать питательные среды и защитить L-формы от осмотического лизиса.
Обычно для L-трансформации используют полутвердые (1,3%) и полужидкие (0,3%) среды. Значительно реже индукция происходит в жидкой среде. Универсальных сред для получения L-форм разных видов бактерий не существует. Состав сред и условия культивирования (температура, длительность, условия аэро- или анаэробиоза, либо культивирование в атмосфере углекислого газа) зависят от вида исходных бактерий.
Для индукции L-форм большое значение имеет состав и концентрация солей, стабилизирующих осмотическое давление среды. Для L-форм грамотрицательных бактерий обычная концентрация NaCl в среде составляет 0,5%, для грамположительных бактерий требуется повышенное содержание веществ, стабилизирующих осмотическое давление. В отдельных случаях I.-формы можно получить на среде с осмотическими стабилизаторами в отсутствии L-трансформирующего фактора.
Универсальным фактором индукции L-форм является пенициллин. Другие антибиотики оказывают избирательное трансформирующее действие в зависимости от видовой принадлежности микроорганизма. Так, бацитрацин вызывает индукцию L-форм стрептококков. Бацитрацин, ванкомицин и ристоциетин индуцируют образование L-форм N. meningitidis.Тетрациклин, канамицин и хлорамфеникол не вызывают образование L- форм.
Индукцию L-форм могут вызвать лизин и лизоцим, антисыворотка и комплемент, некоторые аминокислоты, при этом концентрация аминокислот варьирует в зависимости от вида бактерий.
Сульфаниламиды, соли ртути, кадмия, хрома, лития и многие антисептические вещества, в том числе фенол и формальдегид, не вызывают индукции L-форм. L-формы в небольших количествах могут возникать в популяции как бы спонтанно без видимых внешних воздействий. Предполагается, что трансформация в этих случаях может быть вызвана продуктами метаболизма, накопившимися в среде культивирования.
Основными средами выделения и дальнейшего культивирования L-форм являются триптический перевар мяса, сердечной мышцы крупного рогатого скота, стандартная среда PPLO-агар, экстракт мозга и сердца; в качестве дополнительных факторов роста — дрожжевой гидролизат, печеночный и яичный экстракты. В качестве обязательного компонента используют сыворотку крови лошади или кролика.
Накопленные данные о частоте L-трансформации у разных видов бактерий, однотипность и закономерный характер изменений биосинтетических процессов, морфологических и физиологических признаков, передающихся по наследству, значительно расширяют наши представления об изменчивости бактерий и патогенетическом значении L-форм.
- Читать далее "Некультивируемые формы патогенных бактерий: характеристика, способы выявления"
Редактор: Искандер Милевски. Дата публикации: 8.11.2019