Роль каспаз в апоптозе клетки. Функции каспаз
Центральная роль в исполнении программы апоптоза принадлежит специфическим протеолитическим ферментам каспазам — семейству цистеиновых протеиназ, расщепляющих субстрат в сайтах, следующих за остатками аспарагиновой кислоты (CASPASE — Cysteinyl Aspartat specific proteinase). Вне апоптогенного сигнала каспазы предсуществуют в клетке в форме проэнзимов с незначительной протеолитической активностью.
Наличие готовых к использованию белковых молекул позволяет в нужный момент даже в условиях подавления транскрипции и белкового синтеза быстро подготовить их для выполнения программы апоптоза. Несмотря на различное предназначение, все каспазы обладают значительной структурной гомологией и активируются по одной схеме: малоактивный проэнзим, состоящий из одной полипептидной цепи, подвергается протеолитическому процессингу, в результате которого от молекулы отщепляется N-концевой регуляторный домен, а оставшаяся часть разрезается на две неравные субъединицы, образующие функционально активный гетеродимер; два гетеродимера объединяются в тетрамер активного фермента.
Было бы неверным полагать, что каспазы существуют в клетке исключительно в латентном состоянии в ожидании сигнала к исполнению суицидного акта. Функции активных каспаз в клетке не ограничены исключительно исполнением программы гибели: каспазы-1, -4, -5 и -13 (нумерация соответствует хронологии открытия отдельных ферментов) принимают участие в биосинтезе цитокинов, т. е. участвуют в процессах межклеточной сигнализации.
Например, прежнее наименование каспазы-1 — ICE прямо свидетельствует об этой роли фермента — ИЛ-1-b конвертаза (Interleukin-1b converting enzyme).
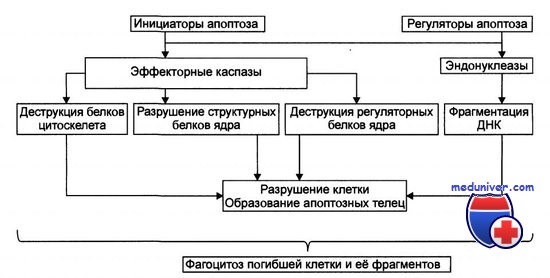
Непосредственно с реализацией программы клеточной гибели связывают действие нескольких ферментов класса каспаз. Принято выделять инициирующие (каспазы-2, -8, -9, -10, -12), участвующие в трансдукции сигнала апоптоза путем трансактивации «нижестоящих» каспаз, и собственно «нижестоящие» эффекторные (или «казнящие») ферменты (каспазы-3, -6, -7), активация которых означает наступление финальной и быстрой стадии «исполнения приговора»: субстратами эффекторных каспаз (в настоящее время известно более 80 таких субстратов) являются жизненно важные для клетки белки — ICAD, PARP, топоизомераза-I, ламины ядерной стенки, микрофиламенты цитоскелета, антиапоптозные белки семейства Вс1-2, гистон HI и др..
Так, в результате деградации PARP [poly-(ADP-ribose)polymerase] прекращается репаративный синтез ДНК; гидролиз гистона HI открывает доступ к межнуклеосомным участкам ДНК для ДНК-фрагментирующего фактора DFF (он же CAD — Caspase-Activated DNase) при каспазазависимом разрушении ICAD (Inhibitor of CAD). Результатом межнуклеосомной фрагментации ДНК является формирование «лестницы ДНК» на электрофореграммах — характерный биохимический признак апоптоза.
Финальной стадии «исполнения приговора» предшествуют минимум две стадии развития событий: акцепция сигнала апоптоза (апоптогенного стимула) и анализ его на соответствие прочим сигналам, в том числе сигналам выживания, совокупность которых часто называют «клеточным контекстом».
- Читать "Рецепторы смерти клетки при апоптозе. Функции рецепторов смерти"
Оглавление темы "Апоптоз клеток":- Роль каспаз в апоптозе клетки. Функции каспаз
- Рецепторы смерти клетки при апоптозе. Функции рецепторов смерти
- Роль митохондрий в апоптозе (гибели) клеток
- Роль факторов роста в апоптозе (гибели) клеток
- Роль гранзимов в апоптозе (гибели) клеток
- Белки теплового шока (HSPs — heat-shock proteins) в апоптозе (гибели) клеток
- Транскрипционные факторы NF-кВ и р53 в апоптозе (гибели) клеток
- Протеасомы в апоптозе (гибели) клеток
- Cигнальные системы гибели и выживания клетки при апоптозе
- Программированная гибель клеток (апоптоз) в цитостатической терапии