Регуляция РНК вирусов. Активность РНК вирусов
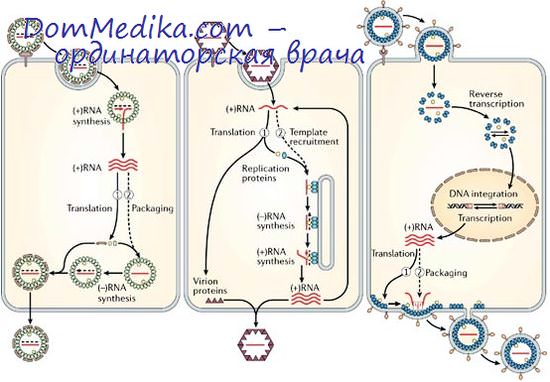
Механизмы, регулирующие транскрипцию, трансляцию и репликацию вирусной РНК, до сих пор изучены недостаточно. В вирусах семейства Picornaviridae единая молекула вмРНК транслируется непосредственно в единый полипептид, и регуляция транскрипции или трансляции отсутствует. Однако в случае вирусов, несущих транскриптазу, у которых синтезируются моноцистронные кмРНК, индивидуальные гены вРНК транскрибируются с различными скоростями и образующиеся в результате молекулы кмРНК транслируются также с различными скоростями.
Количества синтезирующихся молекул разных вирусных белков не эквимолярны и не обратно пропорциональны их молекулярным весам. Более того, относительные скорости синтеза некоторых вирусных полипептидов систематически меняются по мере протекания цикла размножения: неструктурные белки и иногда белки сердцевины часто превалируют на ранних стадиях, капсидные белки — на поздних.
Кроме того, есть основания полагать, что на любой фазе цикла размножения функционирует некий регуляторный механизм, определяющий, как будут использованы молекулы мРНК: для трансляции, для репликации РНК или же — в случае вмРНК— для сборки вириоиов.
Поскольку мы практически ничего не знаем о том, каким образом выполняется такая регуляция в клетках животных, полезно кратко рассмотреть способы регуляции, известные для РНК-фагов (обзоры: Календар, 1970; Стэвис и Огаст, 1970; Козак и Нейтенс, 1972). Возможно, наиболее важное значение имело открытие полифункциональности большинства белков РНК-фагов.
Например, капсидный белок фага f2 является также репрессором синтеза репликазы, а репликаза фага Q подавляет синтез белка, блокируя присоединение рибосом к мРНК. Существуют также указания на то, что «белок созревания» фага f2 остается связанным с вРНК во время ее трансляции и, возможно, репликации. Рибосомы из зараженных бактерий обладают повышенным сродством к вирусной мРНК, и некие «инициирующие факторы», связанные с рибосомами, отбирают для трансляции определенные мРНК.
В регуляции скорости синтеза индивидуальных белков на полицистронных вмРНК важную роль играют вторичная и третичная структуры вмРНК, которые в ходе трансляции определенным образом меняются. Клеточный белок может изменить специфичность полимеразы фага Qp, в результате чего она приобретает способность транскрибировать цепи противоположной полярности. Сходные с таким белком «сигма-факторы» (а) хорошо изучены у ДНК-фагов (Хёрши, 1971).
Эти представления следует принимать во внимание при оценке недавних данных о регуляции выражения РНК-геномов вирусов животных.
1. Богатые аргинином белки сердцевины остаются на протяжении транскрипции и, возможно, трансляции и репликации связанными с вирусной РНК. Хотя имеющиеся в настоящее время данные весьма ограничены, результаты, полученные при изучении вирусов трех различных родов (Orthomyxovirus, Rhabdovlrus и Reovirus), позволяют предположить, что нуклеопротеид нуклеокапсида, обладающий высоким сродством к РНК, играет не только структурную роль.
2. Вирионная транскриптаза, по-видимому, активна лишь тогда, когда она связана с такими нуклеокапсидами, как спиральные рибонуклеопротеиды ортомиксовирусов, парамиксовирусов и рабдовирусов или сердцевина реовирусов. Все еще неясно, являются ли внутренние структурные белки вириона основными субъединицами активного фермента или ферментативная активность присуща минорным белкам вирусной сердцевины.
3. Неструктурный белок вируса гриппа в основном образуется на ранних стадиях цикла размножения и быстро накапливается в ядрышках и на рибосомах. Возможно, это придает рибосомам способность специфически связывать вирусные, а не клеточные мРНК, и таким образом происходит подавление синтеза белка клетки-хозяина.
4. Предшественник капсидных белков полиовируса регулирует трансляцию и репликацию вирусной РНК, а также ее участие в сборке вирионов.
5. Для избирательной транскрипции ранних генов транскриптазой in situ внутри вирусных сердцевин важное значение может иметь конформация вРНК или же возможно, что транскрипция определенных генов до завершения раздевания затруднена топологически.
- Читать далее "Синтез РНК вирусов. Цикл размножения пикорнавирусов"
Оглавление темы "РНК вирусы. Пикорнавирусы":1. Размножение РНК вирусов. Транскрипция и трансляция РНК вирусов
2. Репликация РНК вирусов. Почкование РНК вирусов
3. Регуляция РНК вирусов. Активность РНК вирусов
4. Синтез РНК вирусов. Цикл размножения пикорнавирусов
5. Начальные стадии пикорнавирусных инфекций. Поражения пикорнавирусами
6. Трансляция пикорнавирусов. Этапы трансляции пикорнавирусов
7. Синтез молекул РНК пикорнавирусов. Этапы синтеза РНК пикорнавирусов
8. Регуляция синтеза РНК пикорнавирусов. Ферменты синтеза РНК пикорнавирусов
9. Сборка пикорнавирусов. Этапы сборки пикорнавирусов
10. Регуляция выражения генов пикорнавирусов. Механизмы представления генов пикорнавирусов