Долговременная потенциация как основа нейропластичности - с точки зрения нейрофизиологии
Мы рассмотрели три типа памяти: эксплицитную, имплицитную и эмоциональную память, а также соответствующие нейронные сети. Теперь мы расскажем о том, как меняются нейроны в этих нейронных сетях, обеспечивая консолидацию и сохранение воспоминаний. По общему мнению нейробиологов, изменения происходят в синапсах отчасти из-за того, что именно в области синапсов нейроны влияют друг па друга.
Данная теория была создана в 1928 г., когда испанский анатом Сантьяго Рамон-и-Кахаль (Santiago Ramon у Cajal) предположил, что обучение может вызывать устойчивые морфологические (структурные) изменения эффективности синапсов, возбуждаемых в процессе обучения. Оказалось, что гипотезу Кахаля не так-то легко проверить.
Исследователи по-прежнему сталкиваются с серьезными проблемами, пытаясь проверить гипотезу Кахаля, потому что не уверены, где именно в мозге следует искать синаптические изменения, которые могли бы коррелировать с запоминанием определенных стимулов. Это очень сложная задача.
Представьте себе, будто пытаетесь найти точное местоположение нейронов, отвечающих за хранение имени вашей бабушки. Вы столкнетесь с той же проблемой, если попытаетесь обнаружить нейроны, отвечающие за память о предмете, в мозге обезьяны, выполняющей представленное на рис. ниже задание на зрительное распознавание стимулов.
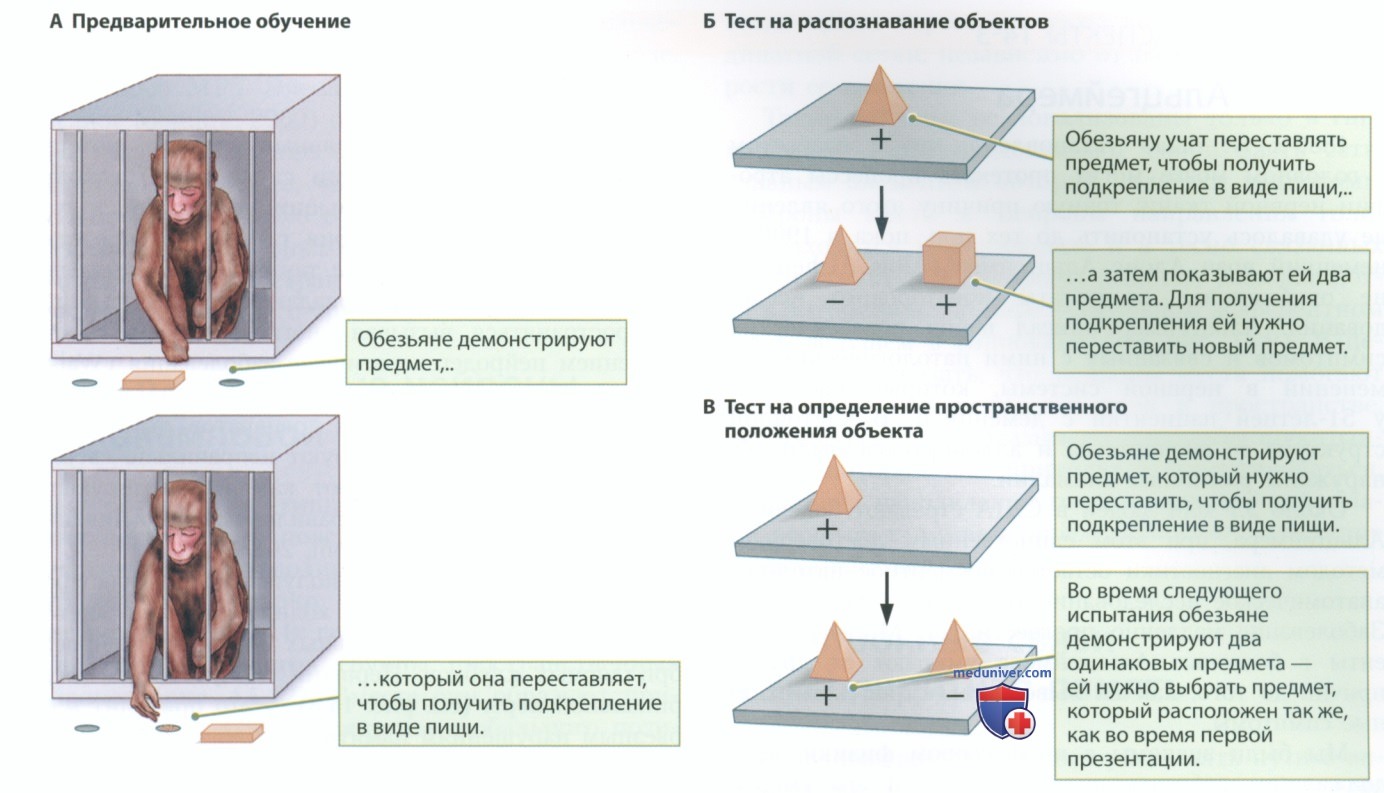
P.S. Нейронная теория Кахаля, которая гласит, что нейрон является элементарной функциональной единицей нервной системы, в наше время признана во всем мире, как и теория о том, что взаимодействие между такими разрозненными клетками обусловливает поведение. Работы Кахаля подробно описаны в отдельной статье на сайте - просим вас пользоваться формой поиска сайта выше.
Один из подходов к поиску нейронных коррелятов памяти предполагает следующее: подтверждение наличия корреляции между синаптическими изменениями в мозге млекопитающего и памятью, локализацию синаптических изменений в специфических нервных путях и анализ самих синаптических изменений. В настоящем разделе приведен обзор исследований, позволивших продемонстрировать, как пережитый опыт коррелирует со связанными с памятью синаптическими изменениями.
Сначала мы рассмотрим стратегию, основанную на физиологии нейронов и пережитом опыте. Затем мы рассмотрим совокупные изменения в нервной системе, коррелирующие с отдельными видами переживаний, которые могут быть как потенциально благоприятными (жизнь в обогащенной среде и обучение выполнению определенных задач), так и потенциально неблагоприятными (постоянное воздействие нейротрофических факторов, гормонов или вызывающих зависимость препаратов). Столь разные переживания вызывают на удивление похожие изменения в общей синаптической организации мозга.
Результаты исследования таких форм поведения, как привыкание (ослабление реакции на стимул) и сенситизация (усиление реакции), на морском зайце (аплизии) показали, что в основе обучения лежат физические изменения синапсов. Способные адаптироваться синапсы мозга млекопитающих участвуют в ассоциативном обучении — формировании временной связи между не связанными друг с другом стимулами путем усвоения того, что стимул А всегда сопровождает стимул Б.
Сформированные ассоциации представляют собой распространенный тип эксплицитных воспоминаний. Примерами таких ассоциаций являются формирующиеся ассоциации между лицом и человеком, запахом и пищей, звуком и музыкальным инструментом. Феномен, лежащий в основе ассоциативного обучения, подразумевает устойчивые изменения в постсинаптическом нейроне после передачи ему возбуждающего сигнала или возбуждающего постсинаптического потенциала (ВПСП) пресинаптическим нейроном.
Относительная простота нейронной организации гиппокампа и простота регистрации постсинаптических потенциалов в гиппокампе делают данную мозговую структуру идеальным объектом для изучения нейронных механизмов ассоциативного обучения. В 1973 г. Тимоти Блисс и Терье Лемо (Timothy Bliss, Terje Lome) продемонстрировали, что многократная электрическая стимуляция идущего в гиппокамп нервного пути обеспечивает постепенное увеличение ВПСП, регистрируемых в клетках гиппокампа. Такое увеличение ВПСП может сохраняться несколько часов, недель или даже дольше.
Блисс и Лемо назвали данное явление долговременной потенциацией (LTP; от англ. long-term potentiation) — она представляет собой сохраняющееся в течение длительного времени повышение эффективности синаптической передачи после высокочастотной стимуляции.
P.S. Постсинаптические потенциалы повышают (возбуждающие/ВПСП) или снижают (тормозные/ТПСП) вероятность формирования потенциала действия (ТПСП — тормозной постсинаптический потенциал).
На рис. 1, А представлен экспериментальный метод исследования долговременной по-тенциации. Он предполагает электростимуляцию пресинаптического нейрона и регистрацию вызванной стимуляцией электрической активности постсинаптического нейрона. Изображение дисплея на рис. 1, А демонстрирует ВПСП, вызванный единичным электрическим импульсом. Стандартные эксперименты предполагают предъявление множественных тестовых стимулов для оценки величины индуцированного ВПСП. После этого применяют сильное импульсное воздействие с частотой в несколько сотен электрических импульсов в секунду (рис. 1, Б).
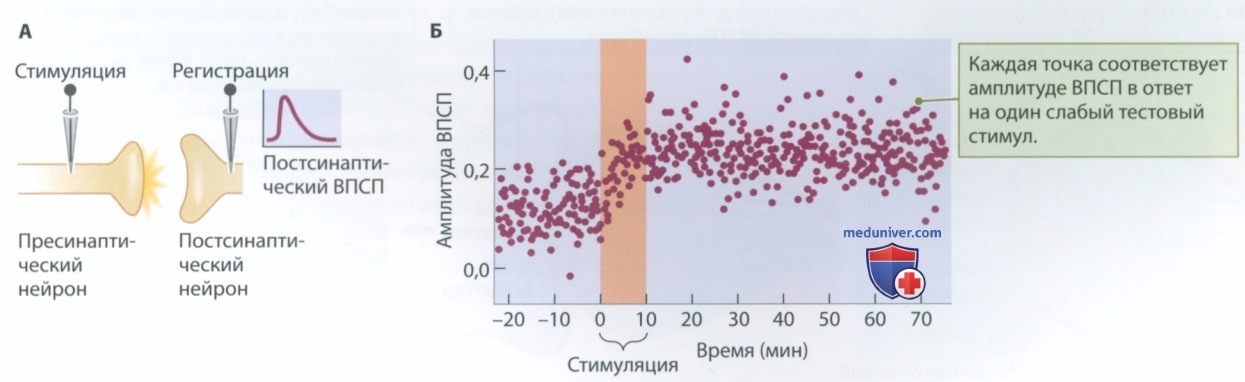
Затем снова воздействуют тестовым импульсом. Увеличенная амплитуда ВПСП сохраняется около 90 мин после высокочастотного воздействия — это и есть долговременная потенциация. Для увеличения ВПСП пресинаптическая мембрана должна выделять больше нейромедиатора или постсинаптическая мембрана должна стать более чувствительной к той же концентрации нейромедиатора (или и то и другое одновременно).
Увеличение выброса нейромедиатора может быть следствием усиления притока ионов кальция в терминаль пресинаптического нейрона, запускающего цепочку событий, которая в конечном итоге приводит к увеличению выброса глутамата. Оно также может быть вызвано выработкой способных к диффузии постсинаптических ретроградных мессенджеров, которые влияют на выброс нейромедиатора. Точный механизм ретроградной сигнальной системы не изучен — ученые предполагают, что функцию ретроградных мессенджеров могут выполнять такие газы, как окись углерода и окись азота. Кроме того, долговременную потенциацию могут модулировать ионотропные и метаботропные рецепторы на терминалях пресинаптического нейрона.
P.S. Применяемая для лечения тяжелой депрессии глубинная стимуляция мозга вызывает изменения, схожие с долговременной потенциацией, — по-видимому, такие изменения стимулируют нейропластичность.
Открытие явления долговременной потенциации привело к революции в понимании процесса сохранения воспоминаний. Когда исследователи изменили воздействие, вызвавшее долговременную потенциацию, они открыли противоположный эффект. Вместо высокочастотной стимуляции (например 100 Гц) они использовали низкочастотную стимуляцию (например, 5 Гц), что привело к уменьшению ВПСП, которое было названо долговременной депрессией (LTD; от англ. long-term depression). Если долговременная потенциация представляет собой механизм формирования воспоминаний, то долговременная депрессия, вероятно, является стирающим старые воспоминания механизмом.
Исходя из того, что долговременная потенциация и долговременная депрессия являются ключом к пониманию лежащих в основе формирования памяти синаптических изменений, можно сделать два предположения. Во-первых, когда животное чему-либо учится, мы можем наблюдать усиление долговременной потенциации в задействованных в обучении нервных путях. Во-вторых, долговременная потенциация должна обеспечивать устойчивые изменения морфологии синапсов, подобные наблюдаемым при формировании памяти. Похоже, что оба предположения верны.
Первоначально исследования долговременной потенциации сводились к изучению возбуждающих глутаматергических синапсов. Выделяемый пресинаптическим нейроном глутамат воздействует на два типа рецепторов на постсинаптической мембране — NMDA- и АМРА-рецепторы (рис. 2, А). АМРА-рецепторы обычно опосредуют ответы, возникающие при высвобождении глутамата из пресинаптической мембраны. Они позволяют ионам натрия (Na+) проникать внутрь клеток, обеспечивая деполяризацию и, следовательно, возбуждение постсинаптической мембраны. Начальная амплитуда ВПСП на рис. 1, А является результатом действия такого АМРА-рецептора.
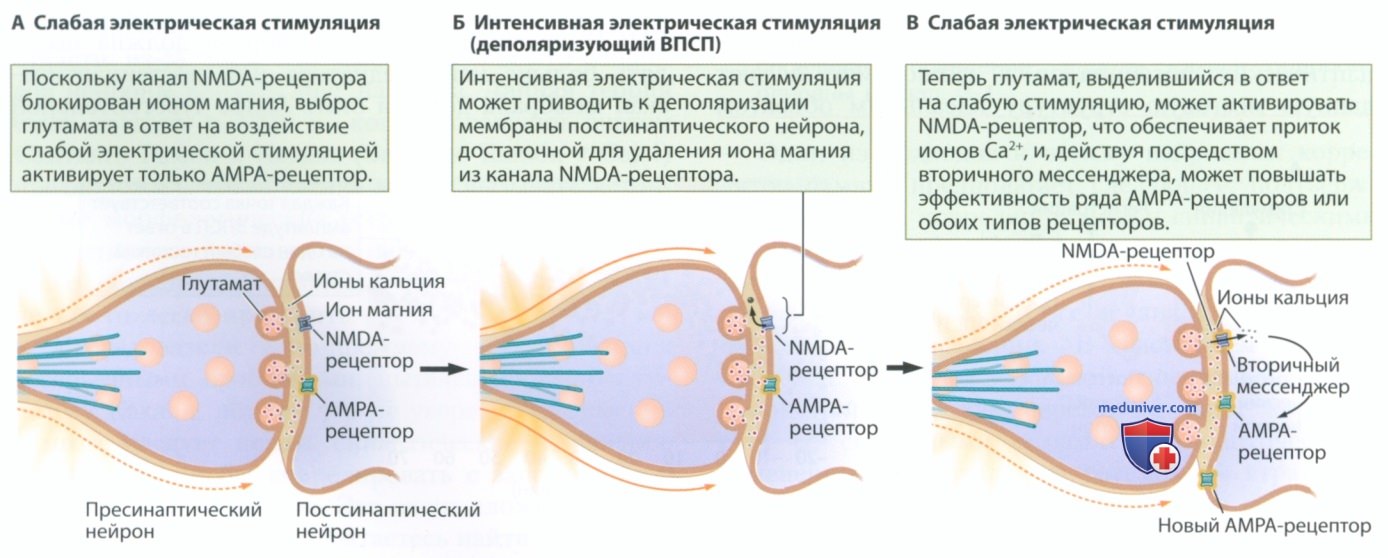
NMDA-рецепторы обычно не реагируют на глутамат, потому что их поры заблокированы ионами магния (Mg2+). NMDA-рецепторы представляют собой ионные каналы с двойным воротным механизмом, которые могут открываться, пропуская ионы кальция, если примерно в одно и то же время происходят два события:
1. Деполяризация постсинаптической мембраны, в результате которой ион магния удаляется из канала NMDA-рецептора (рис. 2, Б). Используемая экспериментаторами сильная электрическая стимуляция представляет собой один из способов удалить ионы магния.
2. Активация NMDA-рецепторов глутаматом, высвобождаемым пресинаптической мембраной (рис. 2, В).
P.S. NMDA — это сокращение от N-метил-D-аспартата; АМРА — это альфа-амино-3-гидрокси-5-метилизо-ксазол-4-пропионовая кислота.
Когда каналы NMDA-рецептора с двойным воротным механизмом открыты, в постсинаптический нейрон поступают ионы кальция, которые воздействуют на вторичные мессенджеры, запуская связанный с долговременной потенциацией каскад событий. К таким событиям относятся повышение восприимчивости АМРА-рецепторов к глутамату, образование новых АМРА-рецепторов и даже ретроградная передача сигналов на пресинаптическую терминаль, усиливающая выделение глутамата. Одно или несколько таких воздействий обеспечивают конечную амплитуду ВПСП, представленную на рис. 1, Б.
Хотя основанные на открытиях Блисса и Лемо исследования сконцентрированы на изучении возбуждающих синапсов, эксперименты на тормозных ГАМКергических вставочных нейронах выявили явления, идентичные с долговременной потенциацией и долговременной депрессией. Эти явления назвали LTPi и LTDi. Это было неожиданное открытие. В то время было принято считать, что тормозным нейронам не свойственна пластичность, однако они оказались пластичными. По-видимому, пластичность ГАМКергических (тормозных) синапсов играет важнейшую роль в регуляции сетей возбуждающих нейронов.
Исследования механизмов долговременной по-тенциации делают очевидной неуверенность нейробиологов в том, где именно локализованы пластические изменения. Наше обсуждение в основном касается изменений постсинаптического аппарата, однако существуют веские аргументы в пользу изменения пресинаптического аппарата (например, MacDougall & Fine, 2014). По-видимому, в целом пластичность обусловлена изменениями на обеих сторонах синапса. Активирующийся первым пресинаптический аппарат может оказаться ключевым элементом на ранних этапах синаптических изменений.
- Вернуться в оглавление раздела "Нейрофизиология"
Редактор: Искандер Милевски. Дата публикации: 2.11.2023
- Два типа памяти - с точки зрения нейрофизиологии
- Какова природа различий между эксплицитной и имплицитной памятью? - с точки зрения нейрофизиологии
- Каковы особенности личных воспоминаний? - с точки зрения нейрофизиологии
- Диссоциация систем памяти - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за эксплицитные воспоминания - с точки зрения нейрофизиологии
- Консолидация эксплицитных воспоминаний - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за имплицитные воспоминания - с точки зрения нейрофизиологии
- Нейронная сеть, отвечающая за эмоциональные (аффективные) воспоминания - с точки зрения нейрофизиологии
- Эволюция систем памяти - с точки зрения нейрофизиологии
- Долговременная потенциация как основа нейропластичности - с точки зрения нейрофизиологии